Dehiscence of the capsule and spore dispersal mechanism is the characteristic features of bryophytes. When the capsule is mature, it degenerates and liberated the spores. Some of the Bryophytes (Riccia, Corsinia) have a simple mechanism for the dehiscence of the capsule and spore dispersal. But others (Funaria, Polytrichum) have special and much more complex mechanisms.
The dehiscence of the capsule and spore dispersal in bryophytes are simple mechanism and special mechanism.
Simple Mechanism
There is no special mechanism for the dehiscence of the capsule and spore dispersal in Riella, Sphaerocarpus, Riccia, Corsinia, and the cleistocarpus mosses (Archidium, Phascum).
Riccia
In Riccia, the jacket layer of the capsule i.e., sporophyte (sporophyte only consists of capsule) and the inner layer of the surrounding venter disintegrate long before the spores mature. Although the spore mass remains surrounded by a venter layer inside the gametophytic tissue of the thallus.
The genus Riccia does not have liberation of spores immediately after they are mature and spores remain within the thallus. Ultimately spores are set free by the progressive death and decay of the surrounding gametophytic tissues.
Sphaerocarpus and Riella
Here, the jacket of a capsule remains intact even after the sporophyte is mature. Spores become free only after the death and the decay of the capsule jacket and the surrounding gametophytic tissues.
Archidium and Phascum
Archidium and Phascum are often called cleistocarpus mosses. In this group of members (sub-class Bryidae i.e., Eubrya of the class Bryopsida), where there is no formation of operculum and peristome.
So, in these mosses, spores are set free by the rotting or irregular splitting of the capsule wall.
Special Mechanism
In other members of bryophytes, there is generally some mechanism for the dispersal of spores. The young sporophyte for the first time being remains enclosed inside the calyptra.
In Hepaticopsida, the seta (where seta is present) elongates rapidly after the spores are formed and the calyptra is ruptured exposing the capsule. In the Bryopsida, the seta elongates gradually, the capsule ruptures after it has been pushed through above the calyptra.
Dehiscence of the Capsule in Hepaticopsida
Marchantiales
In many members of Marchantiales (e.g., Marchantia), the jacket of a capsule at maturity splits longitudinally from the apex to the middle into an indefinite number of segments, but there are no definite predetermined longitudinal lines of dehiscence.
In other Marchantiales (e.g., Reboulia, Cryptomittrium), the upper portion of the capsule jacket comes off like a lid and the lower part remains intact forming a cup.
Jungermanniales
In a great number of Jungermanniales, the mature capsule splits into four valves from apex downwards along predetermined longitudinal lines of dehiscence (e.g., Porella), sometimes those valves become reflexed and hang downwards (e.g., Pellia).
In Ricardia and Metzgeria, the splitting includes the elaterophores. Sometimes the four split valves may remain united at the apex and the spores liberate through four slits between the valves (e.g., Cryptothallus, Pallavicinia).
In Fossombronia, the capsule dehisces irregularly. In many species of Podomitrium, the capsule dehisces by two valves only which remain adherent at the tips. After the dehiscence of the capsule, the spore dispersal in a majority of the Hepaticopsida is helped by elaters which exhibit hygroscopic movement.
Spore Dispersal in Hepaticopsida
According to Ingold (1939), there are three mechanisms affecting spore dispersal among Hepaticopsida. These are–
- The water rupture mechanism is seen among the members of leafy Jungermanniales (Jungermannineae)
- The hygroscopic mechanism is seen in most members of Marchantiales having elaters and in thalloid Jungermanniales (Metzgerineae), and
- The spiral spring mechanism is seen in members of two families of Jungermanniales (Lejeuneaceae and Frullaniaceae).
Dehiscence of Capsule and Spore Dispersal in Other Bryophytes
Anthoceros and Notothylas
In the genus Anthoceros of the class Anthocerotopsida, the capsule dehisces, according to species, by 1 to 4 slits along a predetermined longitudinal line of dehiscence.
In Notothylas, the capsule dehisces by 2 or 4 valves. Finally, spores are shed from the exposed capsule by the hygroscopic movement of valves, pseudo-elaters, and columella.
Andreaea
In some mosses like Andreaea of the sub-class Andreaeidae, the peristome and the operculum are absent in the capsule. When the mature capsule dries, dehiscence takes place along the four longitudinal lines of dehiscence. Thus in most species of Andreaea, the capsule opens by four valves, but these slits extend neither to the apex nor the base of the capsule and the ends of the four valves remain united at the apex.
The opening and closing of the ruptured capsule wall take place by hygroscopic changes. When dry the valves bend outwards -as a result, slits open, and spores are set free, again when wet they (the valves) straighten up and close the slits.
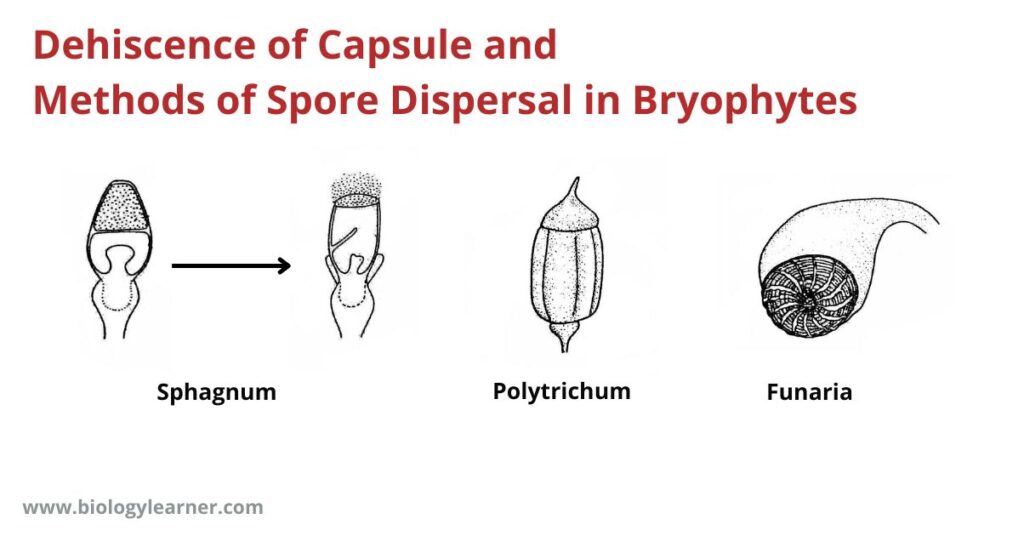
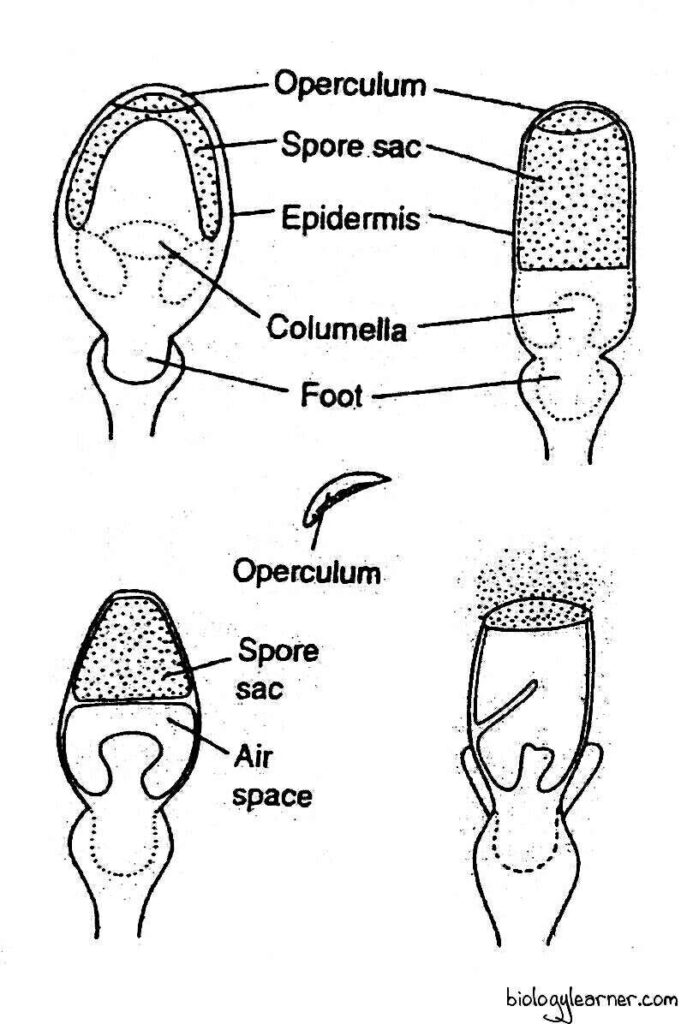
Sphagnum
In Sphagnum (order Sphagnales of the sub-order Sphagnidae), Braunia, and Aulacopllum (order Isobryales of the sub-class Bryidae), the capsule is covered by an operculum, which is forcibly hurled off (due to a pressure created in the columella region beneath the spore-sac) ejecting thereby cloud of dry and powdery spores.
In a large number of Bryidae, The capsule is provided with an operculum and peristome. These two structures together constitute a definite mechanism for the dispersal of spores. The peristome is hygroscopic and this hygroscopic activity of the peristome helps in spore dispersal.
Funaria, Hypnum and Orthotrichum
In some mosses of the sub-class Bryidae like Funaria, Hypnum, and Orthotrichum, the peristome is double layered. The outer peristome is hygroscopic which shows movement according to the presence or absence of moisture and thus helps in the dispersal of spores.
While in absence of moisture, the peristome curves outwards allowing the spores to set free, and in presence of moisture peristome bends inwards preventing the escape of spores. The inner peristome is not hygroscopic and acts only as a sieve.

Polytrichum
In Polytrichum (sub-class Bryidae), the capsule becomes porous. In this genus, the operculum falls off when the mature capsule dries.
Here, the peristome teeth are united with the epiphragm. By drying up the thin-walled cells between the peristome teeth, small openings are formed in the margin of the epiphragm through which the small spores are liberated, a few at a time. As the capsule sways by the velocity of the wind (censer mechanism).
