Polytrichum is one of the largest mosses and a genus of the order Polytrichales.
- It is found in all parts of the world.
- Polytrichum is commonly called hairy cap moss due to the shaggy calyptra enclosing the mature capsule.
| Division: | Bryophyta |
| Class: | Bryopsida |
| Sub-Class: | Bryidae |
| Order: | Polytrichales |
| Family: | Polytrichaceae |
| Genus: | Polytrichum |
Distribution of Polytrichum
- Polytrichum is a prominent genus in the family Polytrichaceae.
- It is represented by about 111 species and is distributed all over the world, mainly in cool temperate and tropical regions.
- Polytrichum mostly prefers to grow in moist and shady places during the rainy season.
- They are found on wet sandy banks of rivers and ponds, on branches of trees under shade (as epiphytes), on rocks and cliffs, and on dry forest bogs.
- Polytrichum juniperinum occurs in extremely dry sites, while P. strictum grows in peat lands. P. sexangulare is found in late-snow areas.
- The species of Polytrichum often form a green carpet of vegetation on moist and shaded walls.
- Some of the common Polytrichum species are P. densifolium, P. alpinum, P. juniperinum, P. xanthopilum, P. strictum, P. formosum, P. sexangulare, etc.
Gametophyte of Polytrichum
The plant body of Polytrichum is gametophytic and consists of an erect leafy shoot.

Morphology of Polytrichum
The plant body is gametophytic. Like Funaria, the gametophyte of Ploytrichum also shows two distinct stages of development: protonema and gametophore. The protonema represents the juvenile stage, while the gametophore is the leafy stage of the adult gametophyte.
Protonema
The protonema is a haploid branched, green filamentous, algal-like structure. It is produced by the germination of the spore. Later, the protonema metamorphoses into the leafy gametophore.
Gametophore
The gametophore i.e., adult gametophyte in Polytrichum is rhizomatous and is differentiated into two parts: the rhizome and the erect leafy shoot.
Rhizome
The rhizome is the horizontally growing underground portion of the gametophore. It bears scale leaves and numerous fluffy rhizoids.
The leaves are small, scale-like, and arranged in three vertical rows (1/3 phyllotaxy). They are usually brown or colourless.
The rhizoids are long, branched, multicellular, thick-walled and characterised by the presence of oblique septa. They are interwoven to form a dense tangled mass.
Rhizoids are colourless and arise from the base of the rhizome.
Rhizoids attach the leafy shoot to the substratum and also help in absorbing water and mineral nutrients from the soil.
Leafy Shoot
The leafy shoot is an erect axis arising from the rhizome. It is the most conspicuous part of the plant and usually grows to a height of about 15-20cm. In P. commune, the shoot is maybe 45 cm in height.
Externally, the leafy shoot is differentiated into a stem-like central axis. The central axis bears two types of leaves: scale leaves and foliage leaves.
The scale leaves are small and produced on the lower portion or transition zone of the central axis. The foliage leaves are comparatively large, dark green, and are arranged spirally on the upper portion of the central axis with a divergence of 3/8.
Each foliage leaf is 6–10 mm long. It consists of two parts; the proximal sheathing leaf base and the diverging narrow limb or blade.
The sheathing leaf base is well-developed, broad, colourless, membranous, and unistratose (one cell thick). It is traversed by a relatively narrow midrib.
The limb is green or brown, lanceolate in outline and has an accuminate apex. It consists of a broad dark green multistratose midrib (nurse of costa) and rudimentary wings. The wings on both sides of the midrib are thin, narrow, and unistratose. The margin of the leaf may be entire (e.g., P. juniperinum) or coarsely toothed (e.g., P. commune).
The erect leafy axis is usually unbranched. Sometimes it is branched and the branch arises from a primordium below the young leaf. The branch primordia occur at intervals both on the rhizome and the erect axis, roughly at the base of every 12th leaf.
The stem grows by means of an apical cell with three cutting faces. It cuts off segments in such a way that the position of the apical cell is shifted in a spiral manner.
Anatomy of Polytrichum
Internal Structure of Rhizome
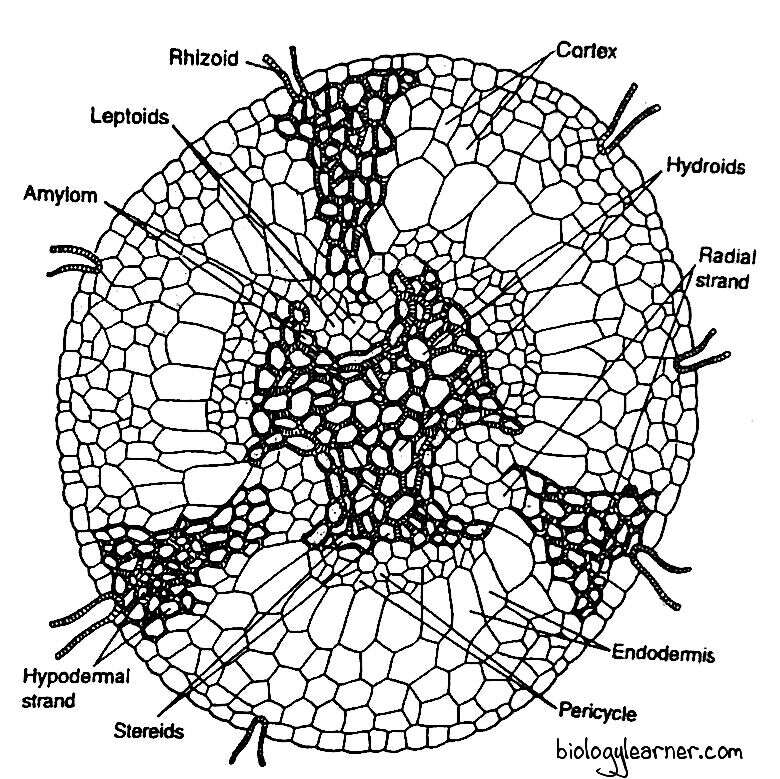
In a cross-section, the rhizome is circular in outline in some Polytrichum species (e.g., P. formusum, P. commune) and triangular with rounded corners in many others. It can be differentiated into the following regions:
Epidermis
The epidermis is the outermost layer of the rhizome. It consists of a single layer of thick-walled cells. The rhizoids, which are the direct outgrowth of the epidermal cells, form a dense tangle.
In the epidermis, stomata are absent.
Cortex
The cortex lies between the epidermis and the central strand. It consists of 3–4 layers of thin-walled parenchymatous cells.
The cortex is interrupted by three hypodermal strands, which extend radially from the periphery towards the centre. Each hypodermal strand is composed of living prosenchymatous cells with a pointed end. Starch grains are present in these prosenchymatous cells.
The hypodermal strands gradually narrow down towards the centre of the rhizome and are connected inward by a group of thin-walled cells. These thin-walled cells, together with hypodermal strands, form a wedge-shaped mass of cells, called radial strands.
Inner to the cortex, there is a layer of large radially elongated cells with suberized thickening on the radial and horizontal walls. This layer can be compared with the endodermis of higher plants. The so-called endodermis is not continuous but is separated by larger cells of radial strands.
Pericycle
In some species of Polytrichum, the pericycle is present between the endodermis and the central cylinder. It is composed of 2-3 layers of thin-walled cells.
The pericycle is not continuous and is absent in the region of furrows, where the centre of the bay is occupied by the lepoids.
Leptoids
In the centre of the bay, a group of polygonal proteinaceous cells is present in each furrow of the central cylinder. These cells are large, elongated, sieve tube-like and are termed the leptoids. The end walls of the leptoids are oblique and, in some cases, the walls are connected to each other through the plasmodesmata.
The leptoids, which more or less resemble the sieve cells of vascular plants, collectively form the leptom. The leptom is the food (photoassimilate) conducting system.
Amylom
On the sideway from the radial strand, each group of leptoids is surrounded by a single layer of parenchymatous cells containing starch. This layer is called the amylom or hydrom sheath.
The amylom thus separates the leptom from the central 3-lobed hydrom.
Central Cylinder
The central cylinder is the central, compact, 3-lobed mass of tissue forming the core of the rhizome. It is made up of two kinds of cells: stereids (or sclereids) and hydroids.
The central mass consists mainly of the stereids. These are thick-walled, elongated, living cells with oblique end walls. The stereids are collectively called the stereom.
The stereom functions as the supporting tissue. In addition, it provides an alternative route for water conduction (Zamski and Trachtenberg, 1979).
Scattered among the stereids are the larger empty cells with very oblique cellulose walls. These are called hydroids.
The hydroids collectively constitute the hydrom. The hydrom is considered to function as the water-conducting tissue.
The stereom and the hydrom together form the hydrom cylinder.
Anatomy of Aerial Stem
A cross-section of the erect aerial stem is more or less circular in outline. Sometimes it is irregular due to the attachment of leaves.
Internally, the erect stem is differentiated into the outermost superficial layer (epidermis), followed by the wide cortex, and the central cylinder.
Epidermis
The outermost superficial layer does not form a clearly defined epidermis. It is single-layered.
Cortex
Within the epidermis is the wide cortex. It is differentiated into the outer and inner cortex.
The outer cortex is thick-walled and composed of compact, elongated prosenchymatous cells that gradually merge into the inner cortex. The inner cortex is made up of loosely arranged, thin-walled parenchymatous cells.
Leaf traces are also seen in the cortical region. Each leaf trace consists of a central patch of colourless, thin-walled, water-conducting hydroid cells (Eschrich and Steiner, 1968). It extends across the cortex and joins the central cylinder.
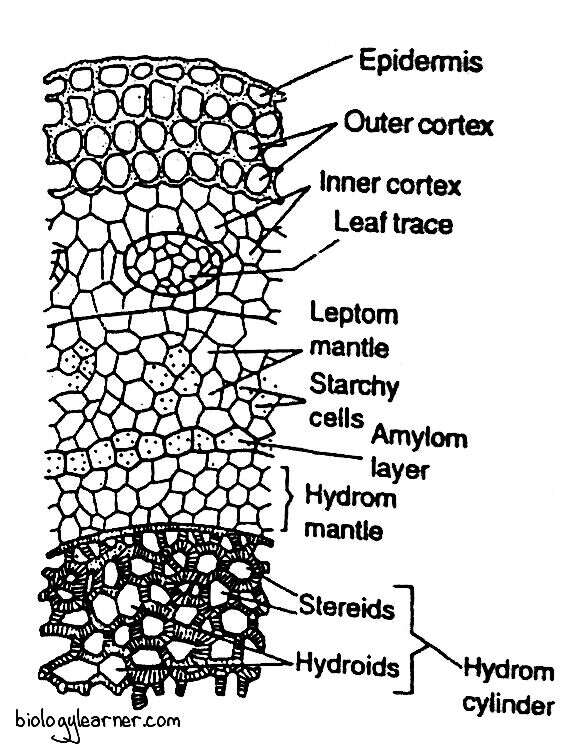
Following the cortex is the endodermis (a layer between the cortex and the leptom, Kawai and Ikeda, 1970). Since the functional aspect of this tissue has not yet been examined, Tranhtenberg and Zamski (1978) suggested that it would be more appropriate to term this layer as the central strand sheath instead of the endodermis. As the cells of this layer bordering the central cylinder contain starch, the endodermis has also been termed the starchy sheath.
Within the endodermis is the rudimentary pericycle, which is not clearly differentiated.
Inner to pericycle is a broad irregular zone composed of thin-walled, sieve tube-like cells. This zone is known as the leptom mantle. It is regarded as equivalent to the phloem of vascular plants.
Internal to the leptom mantle is the hydrom sheath or amylom layer. It is a narrow zone consisting of a single layer, sometimes two layers, of cells containing prominent starch. These cells have dark-brown suberized walls.
Central Cylinder
At the centre of the erect stem is a compact mass of thick-walled cells forming the hydrom cylinder. The hydrom cylinder is composed of two kinds of cells: stereids and hydroids.
The stereids are thick-walled supporting cells constituting the major part of the hydrom cylinder. Interspersed among the stereids are the thin-walled larger empty cells in groups of 2 or 3. These are the hydroids. The hydroids are considered to function as water conduction.
Surrounding the hydrom cylinder is a zone consisting of two or three layers of thin-walled empty (without content) cells. This layer is called the hydrom mantle.
Anatomy of Leaf
Structurally, Polytrichum leaf is the most complex of all the mosses.
In a cross section, the leaf shows a broad midrib flanked by a narrow wing or lamina. The lamina is unistratose (one cell thick) and composed of hyaline cells.
The wide midrib makes up most of the width of the leaf. It is several cells thick (multistratose) in the centre. Gradually, the midrib merges into the wings.
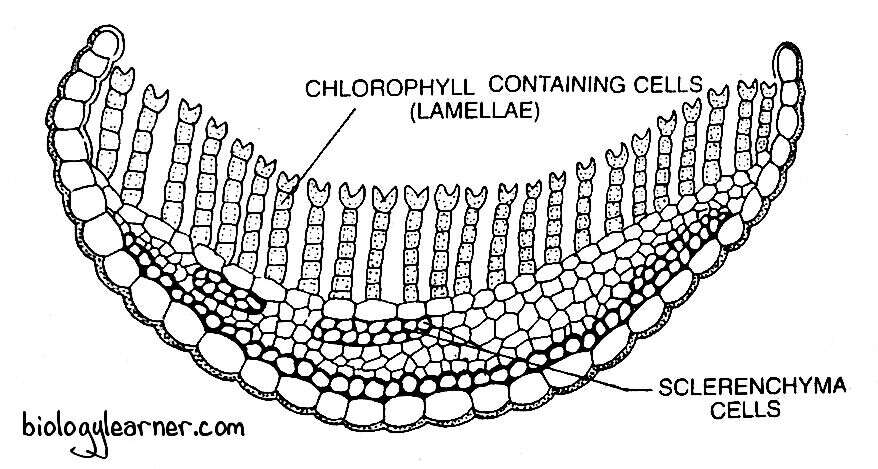
The lower epidermis (epidermis on the abaxial surface) is well developed and consists of a single layer of regularly arranged large cells. These cells have strongly thickened outer walls. Within the lower epidermis, the midrib generally shows one to two, rarely more, layers of a thick band of small sclereid cells with extremely thickened walls and narrow lumina. On the upper side, a narrow and interrupted band of similar slereid cells is present.
Between these two bands of sclereids, the midrib consists of thin-walled parenchyma cells differentiated into a band of large thin-walled parenchyma cells called the deuters and the much narrow empty central parenchyma cells, the hydroids (Hebant, 1971). The deuters lie just beneath the upper band of sclereids. The hydroids help in conducting water, and the deuters are involved in the translocation of assimilates out of the leaf.
On the upper epidermis, there is a layer of large, thin-walled cells from which arise many parallel plates called lamellae. In a cross section, the lamellae appear as one-cell thick (uniseriate) rows of green or dark green cells, standing vertically parallel to each other. Each lamella is made up of 4–7 cells containing chloroplasts.
The leaf lamallae, besides functioning as photosynthetic tissue, also hold water due to capillary force. According to Smith (1971), the lamellae function as pseudomesophyll.
The terminal or distal cells of the lamellae, often referred to as the marginal cells, are slightly larger and colourless. They are variously shaped and are ornamented. In P. juniperinum, the terminal cells are much wider or papillose, but they are bifid in P. commune.
The terminal cells of the adjacent lamellae almost touch each other, thus providing a functional equivalent of the upper epidermis. It is, therefore, termed the upper pseudo-epidermis.
Reproduction in Polytrichum
Polytrichum reproduces by both vegetative and sexual methods.
Vegetative Reproduction
Vegetative reproduction in Polytrichum takes place by the following methods:
Bulbils or Vegetative Buds
Sometimes, vegetative buds or bulbils develop on the rhiziods. Under favourable conditions, bulbils may grow into new plants.
Primary Protonema
The spore, on germination, develops a branched, filamentous structure called the primary protonema. It develops multicellular buds which grow into leafy gametophores.
Secondary Protonema
The protonema that develops from any part of the gametophyte is called the secondary protonema. Each secondary protonema forms buds which grow into gametophores.
Fragmentation of Rhizome
The gametophore in Polytrichum is rhizomatous. The horizontally growing rhizome gives rise to erect, leafy stems at intervals.
Death or breakage of the intervening rhizome results in the development of separate, independent Polytrichum plants.
Apospory
Development of the gametophyte from the sporophyte without the formation of spores is called apospory.
The diploid gametophytic protonema may develop from the unspecialised cells of the various parts of the sporophyte.
Sexual Reproduction
Sexual reproduction in Polytrichum is of the oogamous type. The male gametes (antherozoides) are produced in the antheridium, whereas the female gamete (egg) is developed in the archegonium.
Polytrichum is dioeciuos, i.e., antheridia and archegonium are borne on different gametophores. Sex organs are developed in groups at the tip of the main axis of the gametophore, which is unbranched.
Antheridial Head
The antheridia are borne in a cluster at the tip of the male plant. They are surrounded by a number of bract-like leaves called the perigonial leaves.
The perigonial leaves differ from the vegetative leaves in form and colour and are modified to form a perianth-like structure. Each perigonial leaf is comparatively shorter and usually consists of a broadly expanded sheathing leaf base terminating in a short bristle point. It may be red-brown (P. juniperinum) or dull red (P. piliferum) in colour.
The perigonial leaves lie close together, forming a rosette (perigonial cup) around the antheridia, superficially resembling a small flower.

The antheridia in the perigonial cup occur in small groups at the base of perigonial leaves in the position of lateral buds. The entire antheridial head in Polytrichum is a compound structure, in contrast to that of Funaria. The apical cell of the male gametophore is not used in the formation of antheridia. So, the vegetative axis may grow out through the antheridial head and produce antherida year after year. Such a growth pattern is called “proliferation.”
In a longitudinal section, the antheridial head shows a conscious apical dome surrounded by a number of well developed antheridia. The central dome represents the apical bud containing the growing point (apical cell), which is not used in the formation of antheridia.
Intermingled with the antheridia in the cluster are multicellular hair-like structures called the paraphyses. They are simple filamentous structures consisting of a single row of uniform cells. Some may have spathulate tips.
Antheridium
The male reproductive organ of Polytrichum is the antheridium.
The antheridia are developed in clusters at the apex of the antheridial branch (male gametophore).
Development of Antheridium
- In Polytrichum, the development of antheridium is similar to that of Funaria.
- The antheridium develops from a single embryonic superficial cell at the apex of the male gametophore, called the antheridial initial.
- The antheridial initial enlarges in size, becomes papillate, and protrudes above the neighbouring cells. It divides transversely to form an upper antheridial mother cell and a lower primary stalk cell.
- The primary stalk cell (at a very late stage of antheridium development) undergoes a few divisions to form a few-celled antheridial stalk.
- The antheridial mother cell forms the entire antheridium. It divides by two vertical intersecting walls to form a wedge-shaped apical cell with two cutting faces.
- The apical cell forms segments in two rows in a regular alternate sequence. In this way, 13 to 15 segments are cut off.
- Now the apical cell functions as the operculum cell, which develops the operculum.
- Simultaneously, when the apical cell is dividing, a segment 3–4 cells away from the apical cell starts dividing from the base upward by diagonal vertical walls. The first diagonal wall divides the segment into two cells of unequal size.
- The smaller cell is called the first jacket initial. The larger cell further divides periclinally into an outer jacket initial (second) and an inner primary androgonial cell.
- The jacket initials only divide anticlinally to form a single-layered jacket (wall) of the antheridium.
- The primary androgonial cells undergo further divisions (transversely and vertically) and form the androgonial cells.
- The last generation of androgonial cells is called the androcyte mother cells.
- The androcyte mother cell divides by a diagonal mitotic division and forms two androcytes.
- Each androcyte ultimately metamorphoses into a biflagellate antherozoid, or sperm.

Structure of Antheridium
The mature antheridium of Polytrichum is an elongated structure. It consists of a club-shaped body with a short, few-celled stalk.
The body of the antheridium is encircled by a single-layered jacket. The single-layered sterile jacket surrounds a central mass of androcyte mother cells.
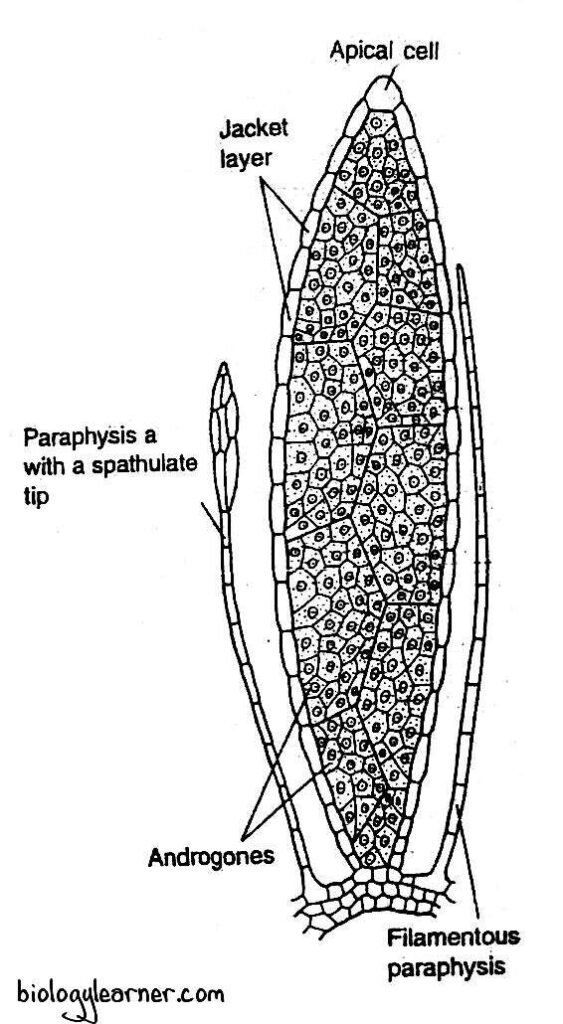
Each androcyte mother cell divides to form two androcytes or spermatids. The androcytes finally metamorphose into antherozoids.
At maturity, a thick-walled operculum (cap) can be seen at the tip of the antheridium. It consists of 4-5 distal tiers of small cells.
Antherozoids
The antherozoid is a unicellular, uninucleate, biflagellated, spirally coiled, elongated structure. The coiled body remains attached to its posterior end with a cytoplasmic vesicle.
Dehiscence of Antheridium
The mature antheridium dehisces with the help of water. The cells of the operculum swell up by absorbing water.
The pressure thus created ruptures the inner wall. Eventually, a narrow pore is formed at the distal end of the antheridium. A large number of biflagellate antherozoids come out through the pore.
Archegonial Head
The archegonia are also borne at the apex of the female plant, like the antheridia of the male plant. They occur in groups. There are usually 3 to 6 archegonia in a group.
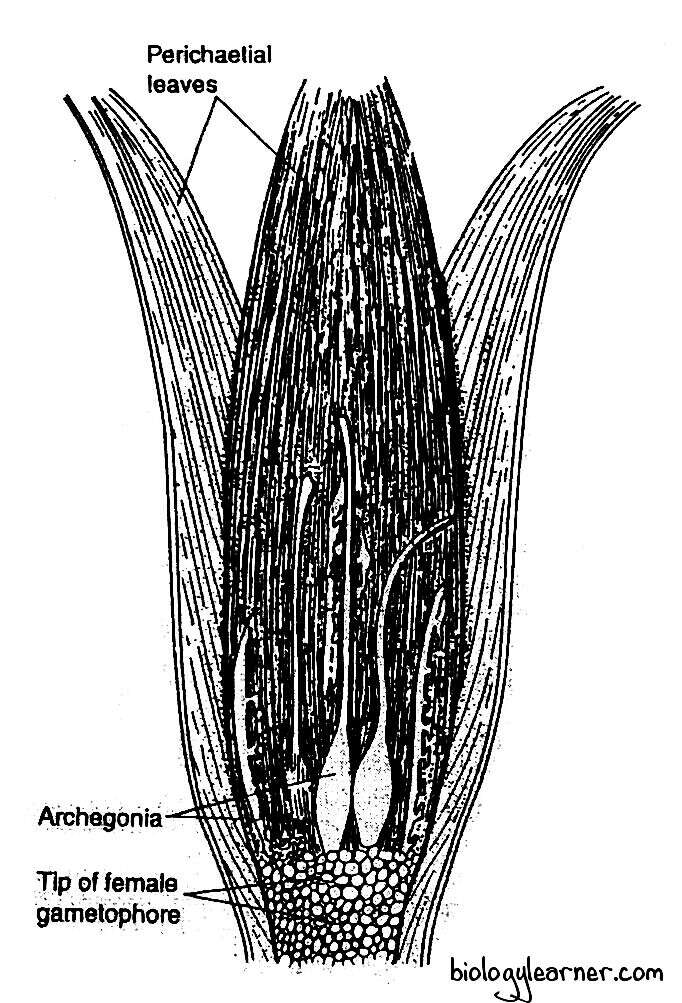
The archegonia intermingled with hair-like multicellular paraphyses in the cluster. The paraphyses are simple filamentous structures and consist of a row of uniform cells.
The group of archegonia is surrounded by perichaetial leaves (foliage-like). The pericheatial leaves overlap at the top of the archegonial cluster to form a closed, bud-like structure called the parichaetium.
Archegonium
The female reproductive organ of Polytrichum is the archegonium.
The archegonia are developed in clusters at the apex of the archegonial branch (female gametophore).
Development of Archegonium
- The archegonium of Polytrichum develops from any apical cell at the tip (growing region) of the female gametophore.
- The apical cell enlarges, slightly projects above the neighbouring cells, and functions as the archegonial initial. It divides by a transverse wall to form a basal primary stalk cell and an upper archegonial mother cell.
- The primary stalk cell undergoes a few irregular divisions and forms the massive stalk of the archegonium.
- The archegonial mother cell divides by three successive intersecting oblique walls, resulting in the formation of three peripheral cells surrounding a tetrahedral axial cell.
- The peripheral cells divide further (transversely, radially, tangentially) to form a 2 or sometimes 3 cells thick jacket or wall of the venter.
- Simultaneously, the axial cell divides transversely to form an outer cell and an inner central cell.
- The central cell divides by an unequal transverse division to form a small venter canal cell and a large egg (oosphere).
- The outer cell functions as an apical cell with four cutting faces (three lateral and one basal).
- The apical cell cuts off successively into three lateral segments and a basal segment.
- Each of the lateral segments divides by a vertical wall, so that the six vertical rows of cells form the single-layered neck of the archegonium.
- The fourth, basal segment forms neck canal cells.

Structure of Archegonium
The mature archegonium of Polytrichum is typically a flask-shaped structure. It has a massive stalk.
Each archegonium consists of an upper, long, twisted neck and a basal, swollen portion, the venter.

The neck is one cell thick and consists of six vertical rows of neck cells, which enclose an axial row of 13 or more neck canal cells.
The venter is composed of a two-cell thick jacket (i.e, wall) of sterile cells. The jacket encircles a small venter canal cell and a large egg.
Fertilization
In Polytrichum, water is essential for fertilization. Due to its heterothallic (dioecious) nature, fertilisation occurs only when the male and female plants grow near each other.
The neck canal cells and the single venter canal cell in the mature archegonium disintegrate to form a mucilage mass. The mucilage imbibes water, swells, and comes out of the archegonial neck by pushing the cap cells apart, forming an open passage leading to the egg known as the neck canal. The mucilage mass contains some chemical substances, such as sugars, that attract the antherozoids.
The rosette-like perigonial leaves function as a shallow splash cup. Rain drops falling on the perigonial leaves can splash the antherozoids up to two feet (Bridie, 1951). This is called the splash cup mechanism.
Splashing drops bring vesicles containing antherozoids to the archegonial cluster. The vesicles dissolve, and the antherozoids are released to swim into archegonia.
Many antherozoids caught in the mucilage enter the archegonial neck due to the chemotactic response and reach up to the egg. Finally, one of the antherozoids fertilises the egg to form a diploid zygote.
Fertilization ends the gametophytic phase.
Sporophyte of Polytrichum
The sporophytic phase in Polytrichum starts with the formation of the diploid zygote after fertilization. The zygote is the first cell of the sporophyte.
Development of Sporophyte
- After fertilization, the zygote immediately secretes a wall around itself and becomes an oospore. It swells up and gradually enlarges in size.
- The zygote or oospore divides first by a transverse wall to form an upper epibasal cell and a lower hypobasal cell.
- Both the hypobasal and epibasal cells divide by two successive intersecting oblique walls to form a young embryo with two growing points at the two opposite ends.
- The epibasal cell produces a wedge-shaped apical cell with two cutting faces. Similarly, the hypobasal cell also produces an apical cell with two cutting faces.
- The entire sporophyte is differentiated by the activity of these two apical cells. Hence, the development of the embryo sporophyte in Polytrichum is bi-apical.
- The apical cell of the epibasal cell develops into the capsule and upper portion of the seta (3/4 portion), while the apical cell of the hypobasal cell forms the slender foot and remaining part of the seta (1/4 portion).
- Both apical cells cut out alternate segments and form the cylindrical young sporophyte.
- The upper region of the young sporophyte divides periclinally to form a four-celled endothecium (a group of central cells), surrounded by eight peripheral cells of the amphithecium.
- The endothecium forms sporogenous tissue and the columella, while the jacket of the capsule develops from the amphithecium.
- Along with the development of the sporophyte, the archegonial wall divides to form a fibrous covering (4-6 cells in thickness) known as the calyptra.
- The calyptra completely encircles the developing embryo (young sporophyte).
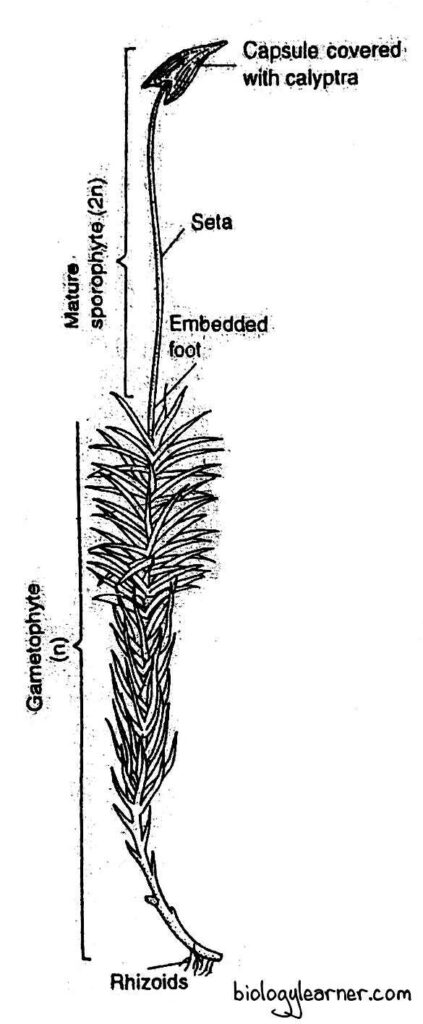
Structure of Sporophyte
The mature sporophyte of Polytrichum is an elongated structure and is relatively complex, like that of Funaria. It is developed at the tip of a female gametophore after fertilization.
The sporophyte is differentiated into three parts: the foot, the seta, and the capsule.
Foot
The basal portion of the sporophyte is the foot. It is a dagger-like conical structure.
The foot is deeply embedded in the apical tissue of the female gametophore and consists of thin-walled parenchymatous cells containing dense cytoplasm. It functions in the anchorage of the sporophyte and in the absorption of water and nutrients from the gametophyte for the developing sporophyte.
Seta
The seta is a long, slender, stalk-like structure that connects the capsule with the foot. It raises the capsule above the apex of the female gametophore.
In a transverse section, the seta shows a single layer of thick-walled epidermis with a well-developed central conducting strand of thin-walled hydroid cells surrounded by the cortex. The cortex is composed of loosely arranged, thin-walled, green parenchymatous cells.
Within the epidermis, there is the hypodermis, consisting of several layers of very thick-walled brown sclerenchymatous cells.
Seta is mechanical in function. It also helps in the conduction of water and nutrients to the developing capsule.
Capsule
The capsule forms the major and most conspicuous part of the sporophyte in Polytrichum. It is long, erect, and generally angular-shaped, but shows a polygonal outline in cross section.
The capsule is developed at the apex of the seta and is almost completely covered by the calyptra, with a hairy surface.

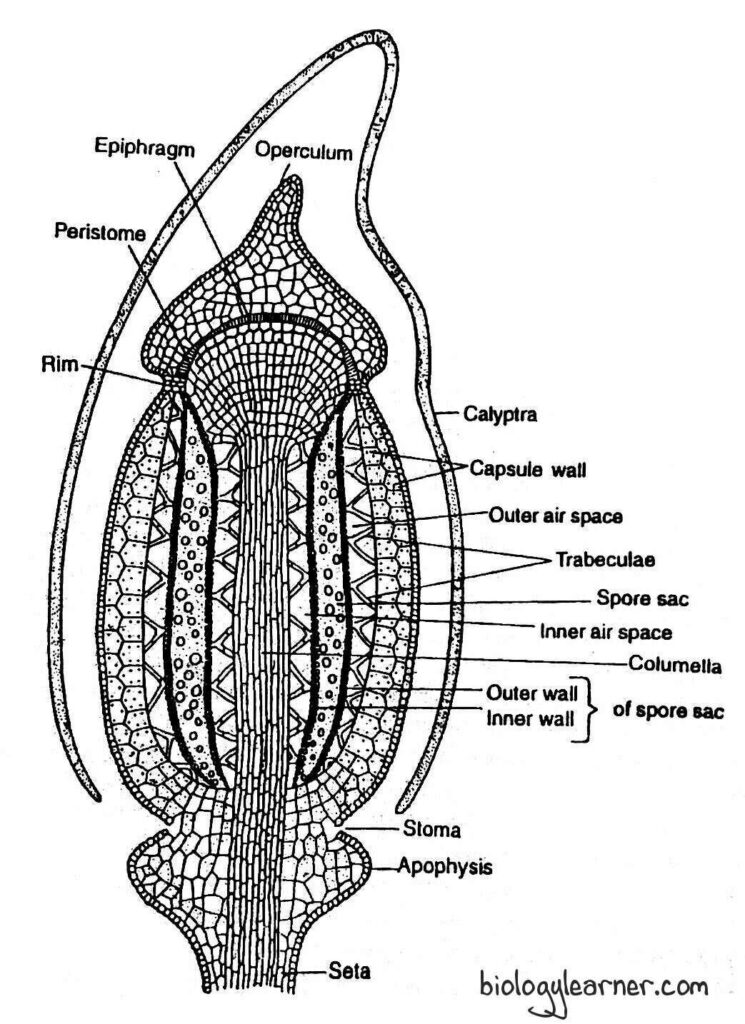
The capsule is differentiated internally into three distinct regions: the apophysis (sterile basal region), the theca (central fertile region), and the operculum or lid (apical region).
Apophysis
The apophysis is the lowermost part of the capsule (also called the neck). It is a bulbous, sterile structure that connects the capsule with the seta below.
Structurally, the apophysis is composed of a single-layered, thick-walled epidermis, which is interrupted by stomata. The sotmata consist of a single ring of guard cells. Usually, two guard cells are present in each stoma.
Below the epidermis is the chlorenchyma, consisting of many thin-walled, loosely arranged parenchymatous cells. These parenchymatous cells contain chloroplasts. Thus, the sporophyte of Polytrichum is actively photosynthetic in function. Therefore, it is not fully dependent on the gametophyte for nutrition.
There is no hypodermis in the apophysis region. The central part of the apophysis is composed of numerous thin-walled cells, forming a conducting strand.

Theca
The theca is the middle, fertile region of the capsule. It lies between the apophysis and the operculum.
A longitudinal section of the theca shows the following regions:
Capsule Wall
The capsule wall, or jacket, is several layers thick and composed of parenchymatous cells.
The outermost layer of the capsule wall is the single-layered epidermis. It consists of compactly arranged cells with thick outer walls. The epidermis of the wall lacks stomata.
The epidermis is followed by typically two layers of spongy parenchyma. The cells of the spongy parenchyma are thin-walled and chlorophyllous. These cells are capable of manufacturing their own food.
Spore Sacs
The spore sacs are situated below the outer air cavities on either side of the columella. Each spore sac has an outer wall and an inner wall. Both walls are made up of two thin-walled cell layers.
The spore sacs are derived from the single-layered archesporium (sporogenous tissue).
The archesporium first develops 4-6 layers of sporogenous cells. The sporogenous cells divide to form many spore mother cells, which fill the cavity (a space between the outer wall and the inner wall) of the spore sac. Later, the spore mother cells divide by meiotic divisions and form haploid spores.
The spore sac in Polytrichum is surrounded by a double air space system, as the air cavities are found on both its outer and inner sides.
Air Cavities
The air cavities (air spaces) are cylindrical and are present on both sides of the spore sacs. The outer air spaces are present just below the spongy parenchyma and outside the spore sacs, while the inner air spaces are found below the spore sacs.
These air spaces are traversed radially by delicate strands (short filaments) of elongated green cells called trabeculae.
The trabeculae of the outer air spaces connect the innermost layer of the capsule wall with the outer wall of the spore sac, whereas those of the inner air spaces connect the spore sac with the central columella.
Columella
The columella is the central part of the theca region. It is made up of compactly arranged parenchymatous cells.
The columella is continuous with the central strand of the apophysis below and extends up to the epiphragm of the operculum above.
Columella helps in the conduction of water and mineral nutrients.
Operculum or Lid
The operculum is the apical region of the capsule. It is a conical structure with a beak or rostrum that forms a cap or lid at the apex of the theca.
The operculum is differentiated from the theca region by a well-marked, narrow circular constriction. At the base of the constriction, there is a diaphragm or rim composed of two to three layers of radially elongated, thick-walled cells. The annulus is absent in Polytrichum.
The columella of the theca is continuous into the operculum. It expands to become a fan-shaped, pale membranous structure known as the epiphragm. The epiphragm stretches like the tympanum of a drum across the mouth of the theca. So the epiphragm is also called the tympanum.
Just within the mouth of the capsule and under the epiphragm, the peristome is present. It is attached below the edge of the diaphragm.
The peristome consists of a ring of 64 short, stout, pyramidal teeth known as peristomial teeth.
Each peristomial tooth is solid and made up of several curved, crescent-shaped, fibre-like cells. Hence, the peristome in Polytrichum is termed the nematodantous peristome.
At maturity, the teeth are joined to the margin of the epiphragm by their tips.
The peristome teeth are not hygroscopic but may control the dispersal of the spores.
Dehiscence of the Capsule and Dispersal of Spores
When the capsule matures, the calyptra falls off. The exposed capsule begins to dry up.
Simultaneously, the operculum shrivels and becomes loose. It eventually drops off, exposing the peristome.
The central tissue (non-sporogenous tissue) of the theca region, except the spores, degenerates at maturity. The spores come to lie in the centre of the capsule.
Meanwhile, the thin-walled cells between the peristome teeth also dry up. As a result, small openings are formed in the margin of the epiphragm.
The spores are dispersed, a few at a time, through these openings as the capsule sways in the wind (censer mechanism).
Spore
The spores are haploid, uninucleate, and yellow in colour. They are very small, 0.010 to 0.011 μm in diameter.
Each spore is surrounded by two wall layers: the outer exosporium (exine) and the inner endosporium (intine). The exine is thick and ornamented, while the intine is thin.
A tiny mass of cytoplasm is present inside the spore wall. It consists of a single nucleus, reserve food materials, and a few plastids (2-4). The reserve food is in the form of lipid droplets.
Germination of Spores and Formation of the Young Gametophyte
The spore is the first cell of the gametophytic generation.
Each spore of Polytrichum germinates upon falling on a suitable, moist substratum. It absorbs moisture from the substratum and increases in size considerably.
The exosporium ruptures at the centre of the aperture region, and the endosporium comes out in the form of a small germ tube.
The germ tube elongates, becomes septate, and produces a green, algal filament-like protonema.
The protonema forms two types of branches: the rhizoidal branches and the chloronemal branches.
The rhizoidal branches are comparatively narrow, have no chloroplasts, and develop below the substratum. They are colourless and possess oblique septa between the cells.
Rhizoidal branches anchor the protonema in the substratum.
The chloronemal branches are spread over the surface of the substratum. They are green and contain chloroplasts in their cells.
The cells of chloronemal branches have partition walls at right angles to the lateral walls (transverse septa) between them.
The chloronemal branches develop many minute green buds. Each bud grows into a leafy gametophore.
