Funaria, a genus of mosses (a group of non-vascular flowerless plants) in the order Funariales, is widely distributed all over the world.
- It belongs to the family Funariaceae.
- Funaria grows during the rainy season as dense patches in shady and damp places.
| Division: | Bryophyta |
| Class: | Bryopsida |
| Sub-Class: | Bryidae |
| Order: | Funariales |
| Family: | Funariaceae |
| Genus: | Funaria |
Distribution of Funaria
- Funaria is the most prominent genus in the family Funariaceae. It is represented by about 117 species and is found in all parts of the world.
- The species of Funaria are terrestrial mosses and grow as green velvety patches in shady and damp places such as moist walls, burnt soil, and the trunks of trees, during the rainy season.
- Funaria hygrometrica is the most common species and is called cord moss due to its twisted seta.
- Some of the common Funaria species are Funaria hygrometrica, F. flavicans, F. apophysata, F. arctica, etc.
Gametophyte of Funaria
The plant body of Funaria is gametophytic and consists of a slender, erect radial stem covered with small, simple leaf-like structures. The gametophytic plant body is the dominant phase of the life cycle in Funaria.

Morphology of Funaria
The plant body of Funaria is gametophytic. The gametophyte shows two distinct stages of development: Protonema (juvenile stage) and Gametophore (adult form).
Protonema
The protonema is a prostrate, branched, filamentous ephemeral structure. Later, the protonema transforms into the leafy gametophore.
Gametophore
The gametophore (adult gametophyte) is differentiated into three parts: rhizoids, axis or stem, and leaves.
Axis or Stem
The axis is about 1 to 3 cm high. It is slender, erect, redial, and often branched. Each branch is extra axillary, i.e., growing from below a leaf.
Leaves
The leaves are simple, sessile, oblong-ovate with an entire margin and a pointed apex. They are spirally arranged on the aerial branches and are more crowded near the apex, forming a rosette.

Each leaf is attached to the stem by a broad base and is traversed by a single distinct midrib. They are borne in 1/3 phyllotaxy, which becomes 3/8 at maturity, and are arranged in three complete spirals.
Rhizoids
The rhizoids are slender, much branched, multicellular and characterised by the presence of oblique septa. They arise from the base of the stem, or axis.
Young rhizoids are colourless but become brown or black as they mature.
The main functions of rhizoids are to attach the axis to the substratum and to absorb water and minerals from the soil.
Anatomy of Funaria
Internal Structure of Axis or Stem
In a transverse section, the mature stem of Funaria shows three distinct regions: the epidermis, the cortex, and the central conducting strand or central cylinder.

Epidermis
The epidermis is the outermost layer of the stem. It is generally one cell thick and consists of small thick-walled tangentially elongated chlorophyll-bearing cells.
Cuticles and stomata or pores are absent in the epidermis.
Cortex
The cortex lies in between the epidermis and the central cylinder. It is many-layer in thickness and composed of large parenchymatous cells.
In the younger part of the stem, the cells of the cortex contain chloroplasts. However, they are lacking in the older section.
As the stem matures, the outer cells of the cortex become thick-walled and are reddish-brown in colour. While the inner cells of the cortex are thin-walled.
The cortex also contains isolated leaf traces (patches of small cells) in the peripheral region.
Central Conducting Strand or Central Cylinder
The central conducting strand or central cylinder is made up of long, narrow, thin-walled colourless cells. These cells lack protoplasm and are called hydroids (dead cells).
The hydroids possibly help in the upward conduction of water and mineral nutrients.
Leaf Anatomy
In a transverse section, the leaf of Funaria shows a well-developed midrib with two lateral lamina, or wings.
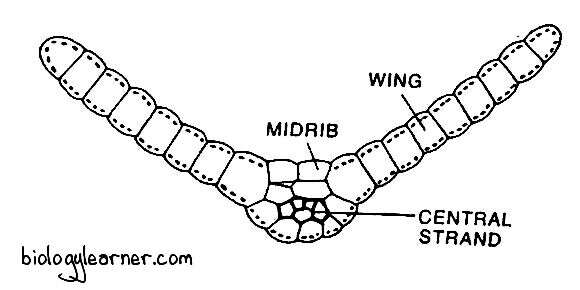
The midrib is several cells thick. It has a small central strand of slightly narrow, thick-walled cells called stereids. They help in conduction.
The leaf lamina is made up of a single layer of parenchymatous polygonal cells. These cells are elongated, thin-walled, and contain large and prominent chloroplasts.
Reproduction in Funaria
Funaria reproduces by vegetative and sexual methods.
Vegetative Reproduction
Vegetative reproduction in Funaria occurs by the following methods:
Fragmentation of Primary Protonema
In Funaria, the spore on germination develops a branched, filamentous structure called the primary protonema.
Under certain circumstances, the primary protonema forms specific colourless separation cells by intercalary divisions. As these cells die, they break up the protonema into several fragments.
The fragments develop into new protonema. Each new protonema bears buds that grow into leafy gametophores.
Secondary Protonema
The protonema that develops from any part of the plant other than the germination of the spore is known as secondary protonema. It may develop from any injured part of the gametophyte, such as rhizoids, stems, leaves, or reproductive structures.
Each secondary protonema forms buds and grows into gametophores.
Gemmae Formation
During unfavourable conditions, many multicellular, green structures called gemmae develop from the terminal cells of the protonema. They become slightly reddish brown in colour at maturity.
With the onset of favourable conditions, the gemmae detach from the parent plant body and grow into new plants.
Bulbils
When such gemmae-like structures are developed on the rhizoidal branches, they are called bulbils. These are brown in colour and devoid of chloroplasts.
Under favourable conditions, each bulbil may grow into a new individual.
Apospory
Development of the gametophyte from the sporophyte without the formation of spores is called apospory.
In the case of Funaria, the gametophytic protonema may develop from the unspecialised cells of the various parts of the sporophyte. The gametophytes thus formed are diploid (2n) but they are normal in appearance.
Sexual reproduction in such gametophytes results in the formation of a tetraploid (4n) zygote. The sporophytes developed from the tetraploid zygotes are sterile and are not capable of producing spores.
Sexual Reproduction
Sexual reproduction in Funaria is of the oogamous type. The male gametes, i.e., antherozoids, are produced in the antheridium (male reproductive organ), and the female gamete, i.e., egg, is formed in the archegonium (female reproductive organ).
Funaria is monoecious (sex organs grow on the same plant), but autoicous, i.e., antheridia and archegonium are borne on separate branches of the same gametophyte.

Sex organs are developed on leafy gametophores. The main shoot of the leafy gametophore bears a group of antheridia at the apex and functions as a male branch. While the female branch bears archegonia and develops later as a lateral outgrowth from the base of the male branch. It grows more vigorously and becomes longer than the male branches.
The species of Funaria are protandrous, i.e., the antheridia mature before the archegonia.
Male Branch
The male branch, or antheridiophore, is the main shoot of the Funaria gametophore.
In a longitudinal section, the male branch shows its apex is enlarged and convex-shaped. It bears a group of reddish brown or orange antheridia in the different stages of development.

The antheridia are surrounded by a number of bract-like leaves called the perigonial leaves. These leaves form a rosette at the apex of the male branch. Perigonial leaves are associated with the cluster of antheridia, collectively forming perigonium (Magill, 1990).
Also, the antheridia are intermingled with a large number of long multicellular sterile hair-like structures known as paraphyses.
Each of the paraphyses is composed of 4 to 6 cells, with an upper large globular cell. They have swollen tips and contain chloroplasts.
Paraphyses protect the developing antheridial heads by minimising transpiration. They may be involved in the dehiscence of antheridia. They also store water between them through capillary action and secrete mucilage, which helps in the liberation of antherozoids. Paraphyses are also involved in photosynthesis due to the presence of chloroplasts.
Antheridium
The male reproductive organ of Funaria is the antheridium. The antheridia are developed in clusters at the apex of the antheridial branch (male branch).
Development of Antheridium
- In Funaria, the antheridium develops from a single superficial antheridial cell at the apex of the male branch called the antheridial initial.
- The antheridial initial enlarges, becomes papillate, and slightly projects above. It divides transversely to form a lower basal cell and an upper outer cell.
- The basal cell undergoes a few divisions to form the embedded portion of the antheridial stalk.
- The outer cell forms the entire antheridium and is called the antheridial mother cell. It divides further by successive transverse divisions and forms a short filament of 2 to 3 cells.
- The terminal cell of the filament divides by two vertical intersecting walls, resulting in the formation of a wedge-shaped apical cell with two cutting faces.
- The apical cell forms segments in two rows in a regular alternate sequence. In this way, 5 to 7 segments are cut off.
- Simultaneously, when the apical cell is dividing, a segment 2 to 3 cells away from the apical cell starts dividing from the base, upwards by diagonal vertical walls. The first diagonal wall divides the segment into two cells of unequal size.
- The smaller cell is called the first jacket initial.
- The larger cell further divides periclinally into an outer second jacket initial and an inner large primary androgonial cell.
- In a transverse section, the primary androgonial cell appears as a triangular cell.
- Similar types of divisions occur in all the upper segments except the apical cell. The apical cell develops into the operculum.
- The jacket initials only divide anticlinally to form a single-layered jacket or wall of the antheridium.
- The primary androgonial cells undergo further divisions (transversely and vertically) to form the androgonial mother cells.
- The last generation of the androgonial cells is known as androcyte mother cells.
- The androcyte mother cell divides by a diagonal mitotic division and forms two androcytes or spermatids.
- Each of the androcytes finally metamorphoses into a biflagellate antherozoid or sperm.

Structure of Antheridium
The mature antheridium of Funaria is a red or orange-coloured elongated structure. It has a club-shaped body with a long multicellular stalk.

The body of the antheridium is encircled by a sterile jacket layer of polyhedral flattened cells. The cells of the jacket are green and contain chloroplasts when they are young. At maturity, the cells become orange or reddish brown. The single-layered sterile jacket surrounds a number of androcytes, which metamorphose into antherozoids.
At maturity, a thick-walled, hyaline operculum, or cap, can be seen at the distal end of the antheridium.
Antherozoids
The antherozoid is an elongated, unicellular, uninucleate, biflagellate, and spirally coiled structure.
The pair of flagella are present at the anterior end of each antherozoid.
Dehiscence of Antheridium
The mature antheridium dehisces with the help of water provided by rain or dew drops. The cells of the operculum swell up by absorbing water.
The pressure thus created ruptures the inner wall. Eventually, a narrow pore is formed at the distal end of the antheridium.
Through this pore, the androcytes spread out in the form of a viscous fluid.
Female Branch
The female branch arises laterally at the base of the male branch.
In a longitudinal section, the female branch shows numerous archegonia interspersed with hair-like, long, multicellular paraphyses at its apex. The terminal cell of the paraphysis is not swollen like the paraphysis present in the male branch.
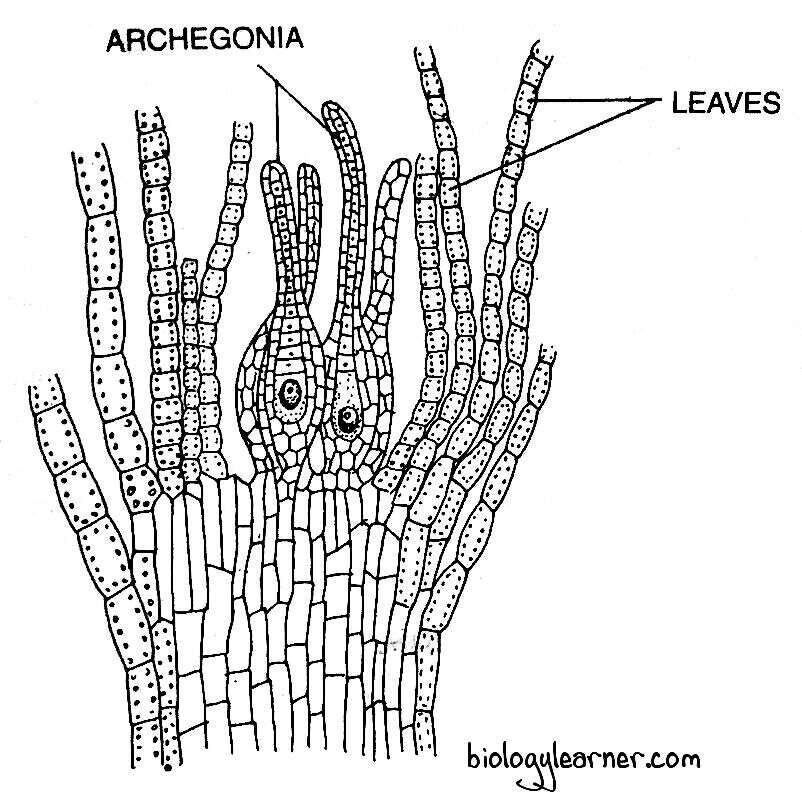
The group of archegonia is enclosed by a number of foliage-like green leaves called the perichaetial leaves. The archegonial cluster with the surrounding perichaetial leaves is known as the perichaetium.
Archegonium
The female reproductive organ of Funaria is the archegonium. Like antheridia, the archegonia are also developed in clusters at the apex of the archegonial branch (female branch).
Development of Archegonium
- The archegonium of Funaria develops from a single superficial cell at the apex of the female branch called the archegonial initial.
- The archegonial initial is enlarged in size. It divides transversely to form the basal cell or stalk cell and a terminal cell (outer cell).
- The basal cell undergoes a few irregular divisions to form the stalk of the archegonium.
- The terminal cell enlarges and functions as an archegonial mother cell. It divides by two intersecting oblique walls and forms an apical cell with two cutting faces.
- The apical cell further divides by three intersecting oblique walls, resulting in the formation of three peripheral cells surrounding a central tetrahedral axial cell.
- The peripheral cells divide anticlinally to form a single-layered jacket or wall of the venter which, by further divisions, becomes two-layered.
- Simultaneously, the axial cell divides transversely to form an outer primary cover cell and an inner central cell.
- The central cell divides into an upper primary neck canal cell and a lower primary venter cell.
- The primary neck canal cell undergoes a series of transverse divisions and forms a row of neck canal cells.
- The primary ventral cell divides by an unequal transverse division to form a small venter canal cell and a large egg or oosphere.
- The primary cover cell functions as an apical cell with four cutting faces (three lateral and one basal). It cuts off successively three lateral segments and a basal segment.
- Each of the lateral segments divides by a vertical wall so that the six rows of cells form the single-layered neck of the archegonium.
- The fourth, basal segment forms neck canal cells.
- In Funaria, the neck canal cells thus have a double origin. The lower and middle neck canal cells in the neck canal are derived from the primary neck canal cell, while those in the upper portion of the neck are derived from the primary cover cell.

Structure of Archegonium
The mature archegonium of Funaria is a much elongated flask-shaped structure. It has a massive long stalk. The stalk attaches the archegonium to the apex of the female branch.
The archegonium consists of an upper elongated slender neck and a basal swollen portion called the venter.
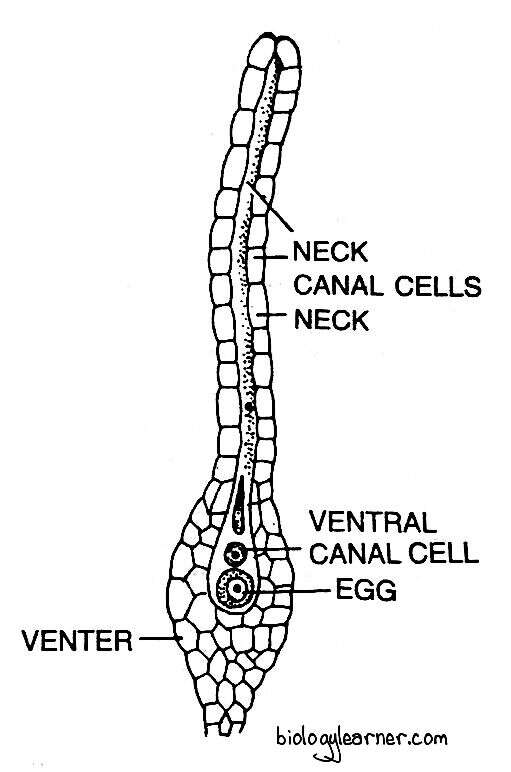
The long neck is slightly tubular, twisted, single-layered and composed of six vertical rows of neck cells, which encircle an axial row of four to ten or more neck canal cells.
The venter is composed of a double-layered jacket or wall of sterile cells. The jacket encloses a small venter canal cell and a large egg or oosphere.
Fertilization
Water is necessary for fertilization in Funaria.
As the archegonium matures, the neck canal cells and the single ventral canal cell disintegrate to form a mucilaginous mass. The mucilage absorbs water accumulated as rain or dew, swells up, and comes out of the archegonial mouth by forcing upon the terminal cover cells, forming a narrow passage down to the egg called the neck canal.
The mucilage mass contains chemical substances (mainly sugars) that attract the antherozoids.
The rosette-like perigonial leaves function as splash cups. Raindrops falling on the perigonial leaves disperse the antherozoids for some distance (raindrops falling on the archegonial cluster situated at a lower level).
A number of antherozoids or sperm caught in the mucilage enter the archegonial neck due to the chemotactic response and reach up to the egg.
Ultimately, only one antherozoid fertilises the egg to form a diploid zygote or oospore.
The gametophytic phase ends after fertilisation.
Sporophyte of Funaria
In Funaria, the sporophytic phase begins with the formation of the diploid zygote after fertilization.
The diploid zygote, or oospore, is the first cell of the sporophyte.
After fertilization, the zygote immediately secretes a wall around itself. It swells up and gradually increases in size.
Development of Sporophyte
The zygote divides first by a transverse wall to form an outer epibasal cell and an inner hypobasal cell.
Both the hypobasal and epibasal cells divide by two intersecting oblique walls to form a young embryo with two growing points at the two opposite ends. The epibasal cell produces an apical cell with two cutting faces, while the hypobasal cell develops an apical cell.
The entire sporophyte is now differentiated by the activity of these two apical cells. Therefore, the development of the embryo sporophyte is bi-apical. The apical cell of the epibasal cell develops into the capsule and upper portion of the seta, while the apical cell of the hypobasal cell forms the foot and the remaining part of the seta.

Both apical cells cut out alternate segments to form the elongated filamentous structure of the young sporophyte.
In a cross-section, the upper portion of the young sporophyte shows two identical segments. The segments divide vertically at right angles to the preceding one to form a four-cell stage, called the quadrant.
Each of the four cells in the quadrant is now divided by an anticlinal wall, resulting in the formation of a smaller triangular cell and a larger rectangular cell.
The four rectangular cells divide by a periclinal division to form a group of four central cells called the endothecium, surrounded by eight peripheral cells of the amphithecium. The layers of endothecium and amphithecium function as fundamental embryonic layers of the sporophyte.
These two groups of cells divide further (anticlinally and periclinally), resulting in the formation of different rings. The amphithecium divides periclinally to form two concentric layers of eight cells each. The inner layer of eight cells is called the first ring. The cells in the first ring divide again and form the outer wall of the spore sac.
The cells of the outer layer divide anticlinally first and form 16 cells. After that, it divides periclinally. The inner part of the 16 cells in this layer is called the second ring. The cells of the second ring later develop the trabeculae.

Again, the cells outer to the second ring divide first anticlinally and then periclinally to form a 32-celled inner third ring. The cells of this layer give rise to the spongy layer.
Similarly, the cells external to the third ring divide periclinally to develop the fourth and fifth rings of 32 cells. The fourth ring forms the hypodermis, while the fifth ring develops the epidermis.
The four cells of the endothecium also divide similarly to the amphithecium. It divides first by a curved vertical wall and then by a periclinal wall. In this way, the endothecium produces four central cells and a ring of eight peripheral cells.
The central cells divide to form the columella. The eight peripheral cells are divided periclinally into an inner layer and an outer layer. The cells of the outer layer divide further to form the archesporium, or sporogenous tissue, while the cells of the inner layer develop the inner wall of the spore sac.
Thus, the archesporium is endothecial in origin. The cells of the archesporium divide and re-divide several times to form two cell layers of thick spore mother cells. The spore mother cells divide by meiotic division and form the tetrad of spores. Elaters are absent in Funaria.
Structure of Sporophyte
The mature sporophyte of Funaria is an elongated structure and relatively complex. It is developed at the apex of a female branch.
The sporophyte of Funaria is differentiated into three distinct parts: the foot, the seta, and the capsule.
Foot
The foot is the basal part of the sporogonium. It is a small, dagger-like conical structure.
The foot is embedded in the apical tissue of the female branch. It helps in the anchorage of the sporophyte and in the absorption of nutrients and water from the gametophyte.
Seta
The seta is a long, slender, stalk-like hygroscopic structure that connects the capsule with the foot. It held the capsule above the apex of the leafy gametophore.
In a transverse section, the seta shows a single-layered epidermis with a central conducting strand of thin-walled cells surrounded by a thick-walled cortex.
Seta provides mechanical support to the capsule. It also helps in the conduction of water and nutrients.
Capsule
The capsule forms the major and most conspicuous part of the Funaria sporophyte. It is a pear-shaped structure and is developed at the apex of the seta.
The capsule is green in colour when young, but becomes bright orange at maturity. The upper portion of the capsule is covered by a cap-like structure called the calyptra.
The capsule can be differentiated internally into three distinct regions: the apophysis (sterile basal region), the theca (central fertile region), and the operculum (apical region).

Apophysis
The apophysis is the basal sterile part of the capsule. It is also known as the neck.
The apophysis connects the capsule with the seta below. It is bounded by the single-layered thick-walled epidermis, which is interrupted by stomata. The stomata are made up of a single ring of guard cells.
Below the epidermis, there is spongy parenchyma. The parenchymatous cells contain many chloroplasts. Thus, the sporophyte of Funaria is actively photosynthetic in function. Hence, it is not fully dependent on the gametophyte for nutrition.
The central part of the apophysis is composed of many elongated, thin-walled, colourless cells forming a conducting strand.
Theca
The theca is the central, slightly bent, fertile region of the capsule. It is situated between the apophysis and the operculum.
In a longitudinal section, the theca shows the following regions:
Capsule Wall
The wall, or jacket, of the capsule, is many layers thick and composed of parenchymatous cells.
The outermost layer of the capsule wall is the epidermis. It is single-layered with very few or no stomata.
A few layers of parenchymatous hypodermis are present below the epidermal layer of the capsule wall. These cells are colourless and are arranged compactly.
Internal to the hypodermis, the cells are loosely arranged and form two to three layers of spongy parenchyma. The cells of spongy parenchyma contain chloroplasts and are capable of manufacturing their own food.
Air Cavities
The air cavities are cylindrical and are present just below the spongy parenchyma and outside the spore sacs.
These cavities are traversed by delicate strands of elongated green cells called trabeculae. The trabeculae connect the innermost layer of the capsule wall with the outer wall of the spore sac.
Spore Sacs
The spore sacs are situated below the air cavity on either side of the columella. They are derived from the single-layered archesporium.
Each spore sac has an outer wall that is 3 to 4 cells thick and an inner wall made up of one layer of small cells.
The cavity (space between the outer wall and the inner wall) of the spore sac is filled with many spore mother cells. The spore mother cells then divide by meiotic divisions and form haploid spores.
Columella
The columella is the central part of the theca region. It is made up of compactly arranged, thin-walled, colourless parenchymatous cells.
The columella is narrow below, connecting the central strand of the apophysis. Its distal part is cone-shaped, which projects into the concavity of the operculum.
Columella helps in the conduction of water and nutrients.
Operculum
The operculum is the apical region of the capsule. It is a very complicated structure and consists of four to five layers of cells.
The operculum is dome-shaped and joins the capsule proper through a notch. At this notch, there is an annular rim or diaphragm composed of two to three layers of radially elongated pitted cells. The diaphragm separates the operculum from the theca proper.
Just above the diaphragm, there is another ring of slightly larger, conspicuous cells, called the annulus. In the annulus, the cells are arranged in five to six superimposed layers. Its two lowermost layers of cells are thin, while the upper cells are thick.

Below the operculum, the peristome is present. It is attached beneath the edge of the diaphragm. The peristome consists of two rings of radially arranged long triangular teeth called peristomal teeth. The peristomal teeth are made up of cuticles and are not cellular in nature.
In each ring of the peristome, there are sixteen teeth. The outer ring of peristome teeth is called the exostome, while the inner ring of teeth is known as the endostome. The exostome is larger, thicker, brown in colour, and ornamented with transverse thickening bands. But the endostome is small, delicate and of a pale colour without transverse bands.

The outer peristome teeth are superposed upon the inner ring. Thus, the peristome proper (whole structure of the peristome) is epicranoid in nature. Inner to peristome teeth lies many thin-walled parenchymatous cells.
The peristome teeth are hygroscopic and help in the dispersal of the spores.
Spore
The spores are haploid, uninucleate, and spherical in shape. They are very small, 0.012 to 0.02 mm in diameter.
Each spore remains surrounded by two wall layers: the outermost exosporium or exine, and the innermost endosporium or intine. The exine is thick, smooth, and brown in colour, while the intine is thin and colourless.
Inside the spore wall, the spore consists of a single nucleus, plastids, reserve food materials, and many oil globules.
Dehiscence of the Capsule and Dispersal of Spores
At maturity, the capsule becomes inverted due to epinasty. It begins to dry up and the thin-walled cells of the annulus break away.
Simultaneously, the operculum shrinks and shrivels. It becomes loose and eventually thrown off. As a result, the peristome teeth are exposed.
The peristome teeth are twisted spirally, appearing like an iris diaphragm. The outer peristome teeth (exostome) are hygroscopic. They show inward or outward movements according to the presence or absence of moisture in the environment. The inner peristome teeth (endostome) do not show any hygroscopic movements but function as a sieve.
In dry weather, the outer peristome teeth lose water and bend outwards with jerky movements. As a result, the slits between the outer teeth become wider. It allows the spores to disperse from the capsule. But only a few spores disperse at a time through the slits due to the inner peristome teeth functioning as a sieve.
In high humidity, the outer peristome teeth absorb water, increase in length, curve inwards, and close the slits. Thus, the exostome prevents the escape of spores in wet weather.
At maturity, the seta also shows jerky movements. The twisting and swinging of seta in dry weather further helps in the dispersal of spores.
Germination of Spores and Formation of the Young Gametophyte in Funaria
The spore is the first cell of the gametophytic generation. Each spore germinates in favourable environmental conditions, e.g., under sufficient moisture.
The exosporium ruptures and the endosporium comes out in the form of one or two germ tubes. The germ tube elongates, becomes septate, and produces a green algal filament-like structure called the primary protonema.
The primary protonema forms two types of branches: the chloronemal branches and the rhizoidal branches.
The chloronemal branches grow horizontally on the moist surface of the soil. They are either erect or remain very close to the substratum. These branches possess conspicuous chloroplasts in their cells and become green in colour. Each chloronemal branch forms the partition walls at right angles to the lateral walls. Most of the branches are of the chloronemal type.
The rhizoidal branches develop below the substratum. They are brown in colour, and the partition walls are oblique to the lateral wall. These branches can produce chlorophyll if exposed to light. Rhizoidal branches anchor the protonema in the substratum.
The chloronemal branches develop many minute green buds behind the cross walls. Each bud grows into a leafy gametophore.
The primary protonema is short-lived. From one primary protonema, many Funaria plants (leafy gametophores) may develop.
According to Sirnoval (1947), the development of protonema under laboratory conditions can be differentiated into two distinct stages: the chloronemal stage and the caulonemal stage.
The chloronemal stage is distinguished by irregular branching, right-angle colourless cross walls, and a large number of discoid chloroplasts that are evenly distributed. These branches never produce buds. After nearly 20 days, the chloronemal stage matures into the caulonemal stage.
The caulonema stage is characterised by regular branching, brown cell walls, oblique cross walls, and relatively fewer chloroplasts. Many buds are formed on the branches of caulonema. The buds later develop into leafy gametophores.
