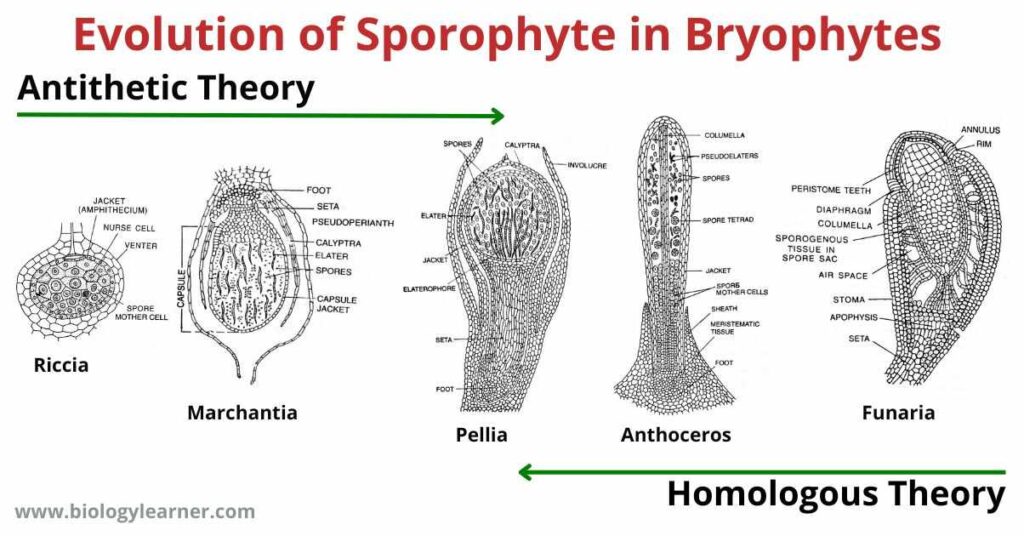
The evolution of sporophyte in Bryophytes have been explained with the help of two theories: one is the Antithetic Theory or Theory of Sterilization and the other one is the Homologous Theory or Reduction Theory.
The antithetic theory states that the evolution of sporophytes has taken place from simple (Riccia, Marchantia) to complex (Funaria, Sphagnum) sporophytes through progressive sterilization of potentially sporogenous tissue.
The other theory (Homologous theory) however assumes that the evolution of sporophyte has taken place from more complex sporophytes (Funaria, Sphagnum) to simpler ones (Riccia, Marchantia) by retrogressive sterilization of sporogenous tissues (exactly opposite is that of antithetic theory).
Antithetic Theory (Theory of Sterilization)
According to this theory, the gametophytic generation is regarded as the original generation, whereas the sporophytic generation was a new phase derived from the progressive elaboration of the zygote of some algal ancestor. which in the course of evolution intercalated into the life cycle between the successive events of fertilization and meiosis and is thus structurally different from the gametophyte.
In 1874, Chelakovsky has first proposed the Antithetic theory which was later on supported by Bower (1890, 1908), Strassburger (1894), Cavers (1910), Chamberlain (1935), and Campbell (1940).
According to Antithetic theory which supports the view of progressive sterilization of potentially sporogenous tissue, the primitive sporophyte is simple with all the cells sporogenous in nature. Gradual sterilization of sporogenous tissues took place.
So that the cells could perform various other functions like providing anchorage, helping in absorption, manufacturing food, spore dispersal, storage of soluble substances and water apart from the usual function of only forming spores and helping in propagation.
First Stage
In Riccia the sporophyte is simple and primitive in nature. There is a single sterile jacket enclosing the central mass of fertile sporogenous cells which divide to form spore-tetrads. The spherical sporophyte is devoid of only foot and seta and remains embedded in the gametophytic thallus.

In Riccia crystallina, a slightly evolved sporophyte is seen. Here some of the potentially sporogenous cells form the sterile nurse cells or nutritive cells.
Second Stage
In Corsinia, further sterilization is noticed, where the basal part of the sporophyte becomes sterile to form a few celled sterile foot. Few sterile nurse cells are however formed from some sporogenous cells.
Third Stage
In Sphaerocarpus, the sporophyte is differentiated into a small bulbous sterile foot, two-celled wide seta, and a capsule. Some of the potential sporogenous cells form the sterile nurse cells. The nurse cells are green in color and contain chloroplasts which provide nutrition to the developing spores.
The single-layer jacket of the capsule is also formed of sterile cells.
Fourth Stage
In Targionia, sporophyte consists of a broad foot, narrow seta, and a single-layered jacket of a capsule. Half of the sporogenous cells form a large number of spirally thickened elaters whereas the remaining half form spore-tetrads.
Fifth Stage
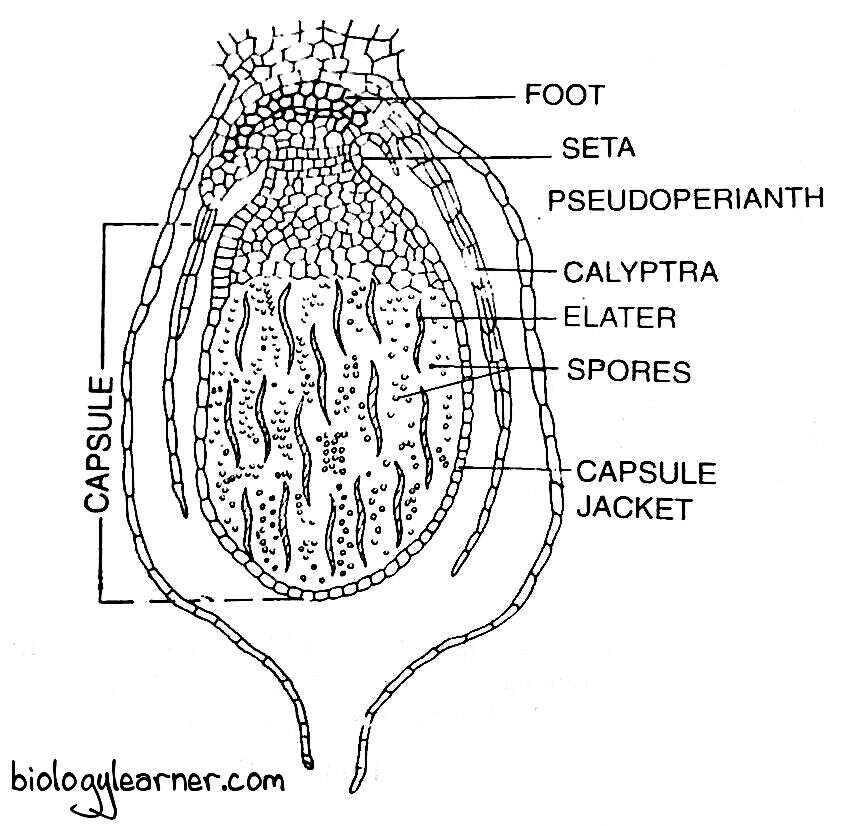
In Marchantia, further sterilization and evolution in sporophyte is seen. The sporophyte is differentiated into a well-developed broad foot, seta, and capsule.
The capsule bears a single-layered sterile jacket and sterile apical cap. Spirally thickened elongated elaters are formed from the potential sporogenous cells.
Sixth Stage
In Jungermanniales (Fossombronia, Riccardia), progressive sterilization of sporogenous tissue can be seen. The sporophyte is differentiated into foot, seta, and a capsule. the capsule consists of the two-many layered jacket. Sporogenous cells form the sterile tissues. The elaterophores form from the sterile tissues which consist of diffuse elaters.
In Pellia, the sporophyte is differentiated into foot, seta and capsule. The capsule contains 2 layers of a sterile jacket. The base of the capsule has a sterile mass of elatophore and some of the potential sporogenous tissues form sterile elaters.
Seventh Stage
In Anthoceros, marked reduction of sporogenous tissues results from increased sterilization. Sporophyte consists of bulbous foot, multilayered capsule, and capsular wall bears functional stomata and chloroplasts.

The sterile central columella is present and sporogenous tissue is in the form of two layers of which few cells produce pseudoelaters. Growth is ensured by the presence of a zone of meristematic tissue. All these characters show the tendency of the sporophyte towards becoming independent.
Eighth Stage
In higher Bryopsida (Musci) like Funaria, and Polytrichum, sterile tissue of complex sporogonium performs diverse functions. Foot, long seta, many-layered capsular walls, columella, wall of spore sac, peristome, operculum, and apophysis are formed of sterile tissues.
In Funaria, the sporophyte is partially independent and is differentiated into well-developed foot, seta and capsule. The sterility of sporogenous tissue is almost maximum. Where the broad apophysis, centrally placed columella, peristome, operculum, many-layered capsular cells, and spore sacs, all are formed of sterile tissues. A small amount of sporogenous tissue forms spores.

The trend of progressive sterilization of potentially sporogenous tissues is evident from the above-stated examples, which is sufficient enough to support the view that the evolution of sporophyte in Bryophytes have taken place from a simpler one to a more complex one, through progressive sterilization of potentially sporogenous tissues.
Homologous Theory (Reduction Theory)
This theory has first proposed by Pringsheim (1876), and supported by Church, Zimmerman, Evans, Fritsch, and Bold.
According to this theory:
- The sporophyte and the gametophyte generations are fundamentally similar in nature and the sporophyte is a direct modification of the gametophyte and is not a new structural type.
- The ancestral Bryophyta had an independent and leafy sporophyte which in the evolutionary course become attached to the gametophyte and gradually has been reduced.
- The complex sporophyte of Funaria is regarded to be primitive.
- Gradual simplification of dehiscence phenomenon.
- Reduction in the photosynthetic tissues and sterile tissues.
- The disappearance of stomata and decrease in capsular wall thickness.
- An increase in potentially fertile sporogenous cells in the path of evolution has given rise to the simple sporophyte of Riccia.
These support the view that the evolution of sporophytes has taken place from more complex sporophytes to simpler ones by retrogressive sterilization of the sporogenous tissue.
