Psilotum is a genus in the order Psilotales. This fern-like vascular plant is mainly found in tropical and subtropical regions.
Psilotum is commonly known as whisk fern due to its whisk-like upward branches and lack of fern.
| Division: | Psilophyta (Pteridophyta) |
| Class: | Psilotopsida |
| Order: | Psilotales |
| Family: | Psilotaceae |
| Genus: | Psilotum |
Distribution of Psilotum
Psilotum is a genus of two species. These two species are well-defined but polymorphic (having two or more different morphs or forms arising from the same genotype).
P. nudum (Syn. P. teiquertum) is very common and widely distributed in tropical and sub-tropical regions of both the eastern and western hemispheres. But P. flaccidum (Syn. P. complanatum) is a rare species found only in tropical islands such as Mexico, Java, Jamaica, Bermuda, etc.
Psilotum is a xerophytic plant, although it grows in various habitats.
The species of Psilotum usually occur on tree ferns and palms as epiphytes. They may also grow terrestrially in soil or in humus-rich crevices of rocks.
P. nudum is a predominantly terrestrial species, while P. flaccidum mainly grows as an epiphyte.
Sporophyte of Psilotum
The plant body of Psilotum is sporophytic.

External Morphology of Psilotum
The sporophytic plant body is differentiated into two parts: a subterranean branched rhizome and an erect aerial stem.
Rhizome
The rhizome is the horizontally growing underground portion of the sporophyte. It is dichotomously branched and brown in color. The branching may be lateral or irregular.
The rhizome contains an endophytic fungus and is covered with many long, multicellar (i.e., 2-3 celled), hair-like rhizoids instead of roots.
Rhizoids anchor the plant body and also help in absorbing water and minerals from the soil.
Aerial Stem
The aerial stem is slender and may be erect (P. nudum) or pendulous (P. flaccidum). It arises from the rhizome. The tip of any branch of the rhizome turns upward and undergoes several dichotomies (apical growth) to form a green aerial shoot.
The aerial shoots are perennial, regularly dichotomously branched, and green in colour. They function as photosynthetic organs.
The basal part of the axis is cylindrical, while the upper green portion may be longitudinally ribbed (e.g., P. nudum) or flattened (e.g., P. flaccidum).
The aerial branches bear numerous small, scale-like appendages, which are often called leaves. The leaves are scattered over these branches.
A few leaves are restricted to the upper portions and bear a group of sporangia (spore-bearing structures) in their axil, which is called the synangium.
Internal Morphology of Psilotum
Internal morphology, or the internal structure of the plant body, is the anatomy of Psilotum.
Anatomy of Aerial Stem
A transverse section of the aerial stem is circular in outline from the base, pentagonal near the first dichotomy, and triangular between successive dichotomies.
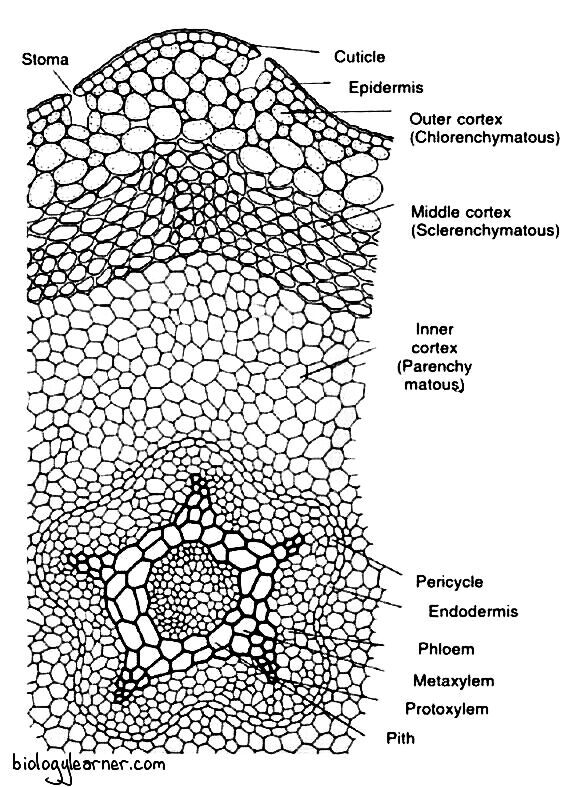
Internally, the stem of Psilotum is differentiated into the outermost epidermis, followed by the extensive cortex and the central stelar region.
Epidermis
The epidermis is the outermost layer of the aerial stem. It is single-layered and consists of closely packed cells. The outer tangential cell walls are heavily cutinized and covered by a layer of cuticle, which is composed of cutin and waxes.
The epidermis is interrupted regularly by stomata. The stomata are sunken in nature and are restricted to the grooves between the longitudinal ridges. They have no subsidiary cells.
Cortex
The extensive cortex is differentiated into three zones: the outer, middle, and inner cortex.
The outer cortex is chlorenchymatous and situates directly beneath the epidermis. It is 2–5 layers thick and consists of loosely arranged, slightly lobed, thin-walled, elongated chlorophyllous cells with intercellular space. The cells of the outer cortex contain starch grains and chloroplasts.
The middle cortex lies internally next to the outer cortex. It is composed of 4-5 layers of vertically elongated, thick-walled sclerenchymatous cells with little or no intracellular spaces. The walls of these cells apparently become lignified in the basal portion of the aerial stem. The cells of the middle cortex provide mechanical support to the stem.
Further inward, there is the inner cortex, which forms the major portion of the stem. It is made up of a few layers of parenchymatous cells without intracellular spaces. These cells show a gradual reduction in the thickness of cell walls towards the centre (i.e., the stelar region). The cells of the inner cortex contain more starch grains.
The cortex is internally delimited by a single-layered endodermis, which consists of vertically elongated cells. These cells have conspicuous casparian strips on the radial and end walls.
Stelar Region
The centre of the aerial stem is occupied by a lobed or flattened cylinder of primary xylem with protoxylem elements at the tip of each lobe. The xylem cylinder may have as many as ten lobes near the transition region from rhizome to aerial stem. The number of lobes is reduced to 5 or 6 in the more distal parts of the stem.
A single-layered pericycle is present just below the endodermis, which consists of thin-walled parenchymatous cells.
The nature of the stele varies in the different portions of the aerial stem. At the extreme base of the stem, the stele is the actinostelic protostele. In the middle part, the stele is siphonostelic, as the centre of the xylem is occupied by a sclerotic pith (a patch of elongated sclerenchymatous cells).
The xylem is exarch (in which protoxylem is directed towards the periphery and metaxylem towards the centre) and composed of only tracheids. The tracheids are scalariform or pitted in the metatoxylem, while the protoxylem has spiral tracheids.
The phloem is internal to the pericycle, lies between the lobes of the xylem, and forms irregular patches. It is composed of sieve tubes and phloem parenchyma.
Anatomy of Rhizome
A transverse section of the rhizome also shows an outermost epidermis, a cortex, and a stelar region.
The epidermis is inconspicuous and consists of thin-walled cells. Rhizoids appear as prolongations of the epidermal cells. Stomata are absent.

The cortex is extensive and divisible into three zones. The outer cortex is parenchymatous and characterised by the presence of intracellular endophytic mycorrhizal fungus.
The middle cortex also consists of parenchymatous cells. But the cells are large and rich in starch grains.
The parenchymatous cells of the inner cortex are often dark brown in colour due to the presence of phlobaphene (an oxidation product of tannins).
The stele is protostelic (haplostelic or actinostelic type) and is surrounded by a distinct endodermis, which is followed by a single-layered pericycle.
The xylem is exarch and completely enclosed by phloem.
Internal Structure of Leaf or Appendage
Structurally, the leaf or appendage of Psilotum is composed of parenchymatous chlorophyllous cells (mesophyll) surrounded by a single layered cutinized epidermis. Stomata are absent in the epidermal layer.

There is no vascular trace in the leaf of P. nudum. But in P. flaccidum, a minute leaf trace appears from the stem, which terminates at the leaf base.
Reproduction in Psilotum
Psilotum shows the alternation of generations of the sporophytic phase and the gametophytic phase. Both sporophytic and gametophytic generations are independent.
The sporophyte of Psilotum reproduces vegetatively as well as asexually. Sexual reproduction occurs in the gametophyte.
Vegetative Reproduction
Vegetative reproduction is usually seen in the sporophyte of Psilotum. Gametophyte also reproduces vegetatively.
In P. nudum, vegetative reproduction occurs through the formation of gemmae or brood bodies (Holloway, 1939; Bierhorst, 1954).
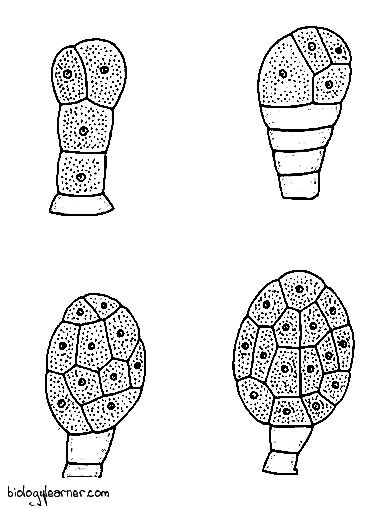
The gemmae are small, multicellular, and oval structures. They are produced in large numbers on the surface of the rhizome in the sporophyte or on the prothallus in the gametophyte. The cells of each gemma are thin-walled and rich in starch grains.
The gemmae of the sporophyte get detached from the rhizome and germinate into new sporophytes. Although the gemmae of gametophyte, after detachment, give rise to new prothallus.
Asexual Reproduction
Asexual reproduction in Psilotum takes place in the sporophytic plant body. It occurs through the formation of spores (i.e., asexual reproductive units).
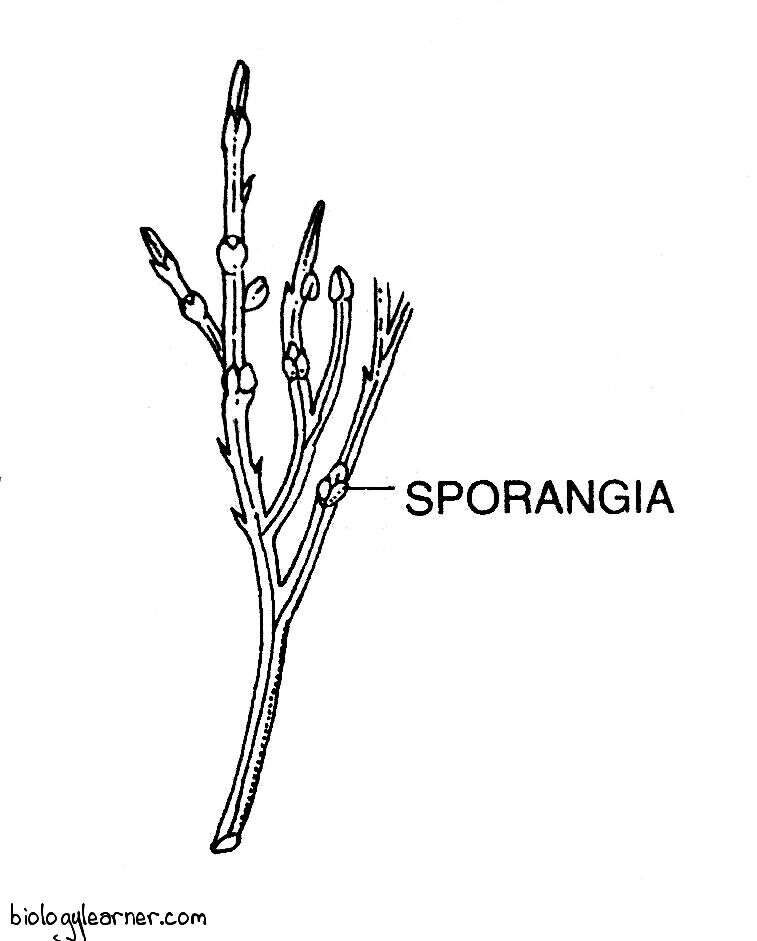
The spores are produced in complex trilobed sporangia called synangium.
Synangium
Synangia are the asexual reproductive organs in Psilotum.
They are borne in the axils of minute bifid leaves on the upper portions of the aerial branches.
Development of Synangium
In P. nudum, the development of each sporangium of the synangium is of the eusporangiate type (arising from several epidermal cells, not from a single cell).
The sporangium develops from a group of superficial initial cells (Bower, 1935). Each initial cell divides periclinally to form an outer primary jacket initial and an inner primary sporogenous cell.

The primary jacket initial undergoes a number of periclinal and anticlinal divisions to form a four to five layered thick wall of the synangium.
Meanwhile, the sporogenous cell divides in various planes and forms a mass of sporogenous cells. Some sporogenous cells later develop into spore mother cells. These cells contain dense, granular cytoplasm. The remaining sporogenous cells gradually degenerate.
Each spore mother cell is diploid and undergoes reduction division (i.e., meiosis) to produce tetrad of haploid spores (four spores).
Structure of Synangium
The mature synangium is a short-stalked, trilobed structure. Each lobe of the synangium corresponds to a single sporangium.
A distinct bilobed, curved foliar appendage is present at the base of the synangium, which surrounds the synangium stalk. The stalk is about 1–2 mm in diameter.

The wall of the synangium is composed of 4–5 layers of cells. The outer wall is thick and consists of relatively long cells. The inner wall separates the three locules, or chambers. Each of the three locules is filled with numerous spores.
Dehiscence of Synangium
At maturity, the synangium dehisces along three vertical lines and liberates the spores.
Spores
Psilotum is homosporous (i.e., it produces spores of one kind) in nature. The spores are of equal size and shape.
Each spore is a bilaterally symmetrical, colourless, kidney-shaped structure. It remains surrounded by two wall layers: the outer exine and the inner intine. The exine is thin and reticulate.
Gametophyte of Psilotum
In Psilotum, the gametophytic phase begins with the germination of spore after asexual reproduction.
The haploid spore is the first cell of the gametophyte.
Germination of Spore and Development of Gametophyte
The spores of Psilotum germinate very slowly. Each spore germinates after four months of falling on a suitable substratum and develops into a gametophyte or prothallus (the gametophyte of pteridophytes is called the prothallus).
Darnell-Smith (1917) was the first to observe the early stages of spore germination and gametophyte development.
The spore germinates exosporically (the gametophyte develops outside of the spore wall). The exine ruptures along the median slit, and the intine comes out in the form of a small tubular outgrowth.

Later, a transverse wall cuts off the outgrowth from the remainder of the spore. The whole structure becomes two-celled.
The upper cell divides obliquely to form an apical cell with three or four cutting faces. The apical cell divides further and forms a mass of thin-walled cells (i.e., the gametophyte).
At this stage of development, the gametophyte becomes infected by an endophytic fungus.
Structure of Gametophyte
The mature gametophyte or prothallus of Psilotum is a subterranean, cylindrical, dichotomously or irregularly branched, elongated structure. Its size usually varies from 0.5 to 2 mm.
The gametophyte is often dark brown in colour due to the presence of an endophytic fungus in the cells. The fungus is probably Cladochytrium tmesepteridis (Beirhorst, 1953). The surface of the gametophyte is densely covered by brownish, hair-like rhizoids.
The gametophyte grows underground as a saprophyte.

Internally, the mature gametophyte is mostly composed of thin-walled parenchymatous cells. The peripheral cells are highly cutinized and are resistant to decay.
Some cells of the outermost layer give rise to rhizoids. A symbiotic fungus is present in many internal cells of the gametophyte.
The centre of the gametophyte also consists of parenchymatous cells, with no trace of any vascular tissues.
Holloway (1939) described certain unusual features of some of the gametophytes of tetraploid P. nudum. Here, these gametophytes show a central vascular strand. The vascular strand is composed of xylem with annular, scalariform, and reticulate tracheids, surrounded by phloem and a clearly recognisable endodermis. There is also a cortex of parenchymatous cells between the vascular strand and the superficial epidermal layer.
Hence, Psilotum is the only plant where the vascular system develops in the gametophytic generation.
Sexual Reproduction
Sexual reproduction in Psilotum takes place in the gametophyte or prothallus. The gametophytes are monoecious i.e., homothallic.
Sex organs are scattered over the surface of the prothallus. The male sex organ is the antheridium, while the female sex organ is the archegonium.
Antheridia and archegonia are both borne in large numbers. But the antheridia are usually more in number than the archegonia.
Antheridium (Development and Structure)
The antheridium is the male sex organ of Psilotum. It develops from a single superficial cell of the prothallus called the antheridial initial.
The antheridial initial divides periclinally to form an outer jacket initial and an inner primary androgonial cell. The jacket initial undergoes repeated anticlinal divisions and forms a single-layered wall of the antheridium.

The primary androgonial cell divides in all the possible planes to form a mass of androgonial cells. At this stage, the antheridium bulges above the surface of the prothallus as a small protuberance.
The last generation of androgonial cells is called the androcytes. Each androcyte finally metamorphoses into a spirally coiled, multiflagellate antherozoid or sperm.
The mature antheridium of Psilotum is more or less spherical in shape. It consists of a single-layered jacket which surrounds many androcytes.

At maturity, a special cell of the antheridial wall called the opercular cell disintegrates and forms a narrow pore (Beirhorst, 1954). Many antherozoids come out through this pore.
The number of antherozoids in the antheridium varies from 32, 64, 128 to 512.
Archegonium (Development and Structure)
The archegonium is the female sex organ of Psilotum. It is also developed from a single superficial cell of the prothallus and is known as the archegonium initial.
The archegonial initial divides periclinally to form an outer primary cover cell and an inner central cell. The primary cover cell divides by two intersecting anticlinal walls at right angles to each other, forming four cells. These four cells function as neck initials. However, a long sectional view at this stage reveals two neck initials and a central cell.

The neck initials undergo several transverse divisions to form a long archegonial neck, arranged in four vertical rows of cells. Each row contains four to six cells.
Simultaneously, the central cell divides transversely into an upper primary neck canal cell and a lower primary ventral cell. The primary neck canal cell divides by a transverse wall and forms two neck canal cells. In most cases, the wall separating the two neck canal nuclei has not been observed. As a result, a binucleate neck canal cell is formed.
The primary ventral cell undergoes an unequal transverse division to form a small ventral canal cell and a large egg.
The archegonium of Psilotum consists of an upper, long, protruding neck and a basal, swollen venter. The venter portion remains embedded in the surface of the prothallus, in direct contact with the surrounding vegetative cells.

The long neck is composed of four vertical rows of cells, and each row consists of four to six cells. It encircles possibly two neck canal cells. The venter contains a ventral canal cell and a single egg cell.
Fertilization
As the archegonium matures, the cell wall of the lower tier of neck cells becomes cutinized, and the upper portion of the neck breaks away.
In the presence of water, the neck canal cells and the single ventral canal cell disintegrate to form a mucilaginous mass. The mucilage absorbs moisture, swells up, and forces open the uppermost tier of neck cells, forming a free passage down to the egg.
The liberated antherozoids swim freely in a thin film of moisture and reach the archegonium. They enter the archegonial neck due to the chemotactic response and reach up to the egg.
One of the antherozoids fuses with the egg and forms a diploid zygote.
Embryo (New Sporophyte)
The zygote is the first cell of the sporophyte.
After fertilization, the zygote increases in size and fills up the cavity of the venter. It divides first by a transverse wall to form an upper epibasal cell (directed towards the neck of the archegonium) and a lower hypobasal cell (directed below).
The epibasal cell gives rise to the sporophytic shoot system (both rhizome and aerial branches), and the hypobasal cell divides in various planes to develop into a bulbous foot of the sporophyte.
As the shoot-forming apical cell is directed towards the archegonial neck, the mode of embryo development (embryogeny) in Psilotum is exoscopic.
With the development of the embryo, a cushion-like gametophytic tissue (so-called calyptra) is formed above the embryo. Later, it is lifted and ruptured by the growing axis of the embryo.
Meanwhile, the foot gives rise to haustoria-like projections. The haustoria absorb nutrients from the gametophyte until the sporophyte becomes physiologically independent.
As soon as the young sporophyte gets infected with the mycorrhizal fungus, it detaches from the gametophyte and becomes independent.

I discovered your blog site internet site on yahoo and appearance some of your posts. Always maintain inside the good operate. Seeking forward to reading more…