Lycopodium is a genus in the order Lycopodiales. This flowerless vascular plant grows mainly in tropical and subtropical forests.
Lycopodium is popularly known as club moss or ground pine due to its moss-like appearance and club-shaped large strobili.
Distribution of Lycopodium
Lycopodium is a genus of about 400 species. It is widely distributed both in tropical and subtropical regions of the world, but more common in tropical zones. Some species are also found in arctic and temperate regions.
Most of the species prefer to grow in moist and shady places rich in humus and other organic compounds.
The species of Lycopodium show various habits. They are either prostrate, semi-erect, or pendent. Species like L. lucidulum and L. reflexum are erect and shurby, while L. volubile is a climber.
A few species, such as L. linifolium and L. phlegmaria, are epiphytes and grow hanging from tree trunks.
- Some of the common Lycopodium species are: L. phlegmaria, L. carinatum, L. clavatum, L. cernuum, L. squarrosum, L. volubile, etc.
Classification of the Genus
The various species of Lycopodium exhibit great diversity in their form, habit, growth, anatomy, etc.
Based on such diversity, Pritzel (1900) divided the genus Lycopodiun into two sub-genera: Urostachya and Rhopalostachya.
In the sub-genus Urostachya (e.g., L. phlegmaria, L. selago), the plant body is erect or pendent, rarely branched, but the branching is always dichotomous. But in the sub-genus Rhopalostachya (e.g., L. clavatum, L. inundatum, and L. cernuum), the plant body is prostrate or semi-erect and profusely branched, showing dichotomy.
Table: Difference between Urostachya and Rhopalostachya
| Characters | Urostachya | Rhopalostachya |
|---|---|---|
| Stem: | Erect or pendent | Prostrate or semi-erect |
| Branching of stem: | Dichotomous | Dichotomous at first, branching becomes monopodial later |
| Roots: | Originate only from the base of the stem | Arise adventitiously all along the length of the prostrate stem |
| Strobilus: | May or may not be organised | Well organised |
| Examples: | L. phlegmaria, L. lucidulum, L. selago, L. compactum, etc. | L. cernuum, L. clavatum, L. densum, etc. |
Sporophyte of Lycopodium
The plant body of Lycopodium is sporophytic.
The sporophyte is herbaceous. It may attain a height of one or two feet.

External Morphology of Lycopodium
The sporophytic plant body of Lycopodium is differentiated into three parts: stem, roots, and leaves.
Stem
The stem is usually weak, slender, and rhizomatous in all the species of Lycopodium.
In the sub-genus Urostachya, the stem is either erect (in terrestrial species) or penden (in epiphytic species) and rarely branched. But the branching is always dichotomous.
In the sub-genus Rhopalostachya, the stem is creeping on or below the soil surface, bearing upright branches.
The branching is dichotomous at the beginning, but later it becomes monopodial.
The stem and branches are thickly covered with many small leaves.
Leaves
The leaves are small, simple, sessile, eligulate, and usually lanceolate in outline with a broad base.
The size of the leaf varies greatly in different species. It is 2–10 mm in L. cernuum. The leaf of L. phlegmaria may be 2–3 cm long.
Usually, the leaves are spirally arranged (e.g., L. annotinum, L. clavatum). But in some cases, they may be arranged in whorls (e.g., L. cernuum, L. verticillatum) or opposite pairs (e.g., L. alpinum). The leaves are all alike in size and shape, i.e., homophyllous. But in a few species, such as L. volubile and L. complanatum, they are heterophyllous, particularly on lateral branches. In L. volubile, the leaves are arranged in four vertical rows on the stem, two smaller and two larger in size.
The leaves of Lycopodium are known as microphylls as they possess a single unbranched median vein that fails to reach the apex.
Generally, the leaves near the apical region of the branches bear sporangia (spore-bearing structures) and are known as sporophylls.
Roots
The mature sporophyte bears only adventitious roots as the primary root is short-lived.
In the sub-genus Urostachya, the roots are developed only from the base of the stem. In some species, such as L. selago and L. pithyoides, the roots arise endogenously from the pericycle, do not penetrate the cortical region of the stem, grow downward through the cortex, and come out only at the base of the stem. A transverse section of the stem at the basal region of these species shows many roots in the cortex. Such roots are called cortical roots or inner roots.
In the sub-genus Rhopalostachya, roots are also adventitious but arise all along the length of the prostrate stem, either singly or in groups.
The branching of roots is dichotomous. In many species, the dichotomy is obscure.
Internal Morphology of Lycopodium
Internal morphology, or the internal structure of the plant body, is the anatomy of Lycopodium.
Anatomy of Stem
A transverse section of the stem is more or less circular in outline. Anatomically, the stem of Lycopodium is differentiated into a superficial epidermis, a broad cortex, and a central stelar region or vascular cylinder.
Epidermis
The epidermis is the outermost superficial layer of the stem. It is one cell in thickness and composed of parenchymatous cells. The outer walls of the cells are thick and cutinized.
The epidermis is regularly interrupted by the presence of stomata.
Cortex
Internal to the epidermis is the wide cortex, which is highly variable in its structure and thickness in different species. The cortex may be homogeneous or divided into three concentric zones (i.e., heterogeneous).
In species where the cortex is homogeneous, it consists of only parenchymatous cells with intracellular spaces (e.g., L. phlegmaria) or only sclerenchymatous cells without any intracellular spaces.
The heterogeneous cortex is found in species like L. clavatum, L. cernuum, etc. In L. clavatum, the outer and inner regions of the cortex are made up of thick-walled sclerenchymatous cells, while the middle region consists of large, thin-walled parenchymatous cells with many intracellular spaces. In L. cernuum, the cortex is differentiated into outer and inner zones of parenchymatous cells and a middle zone of sclerenchymatous cells.
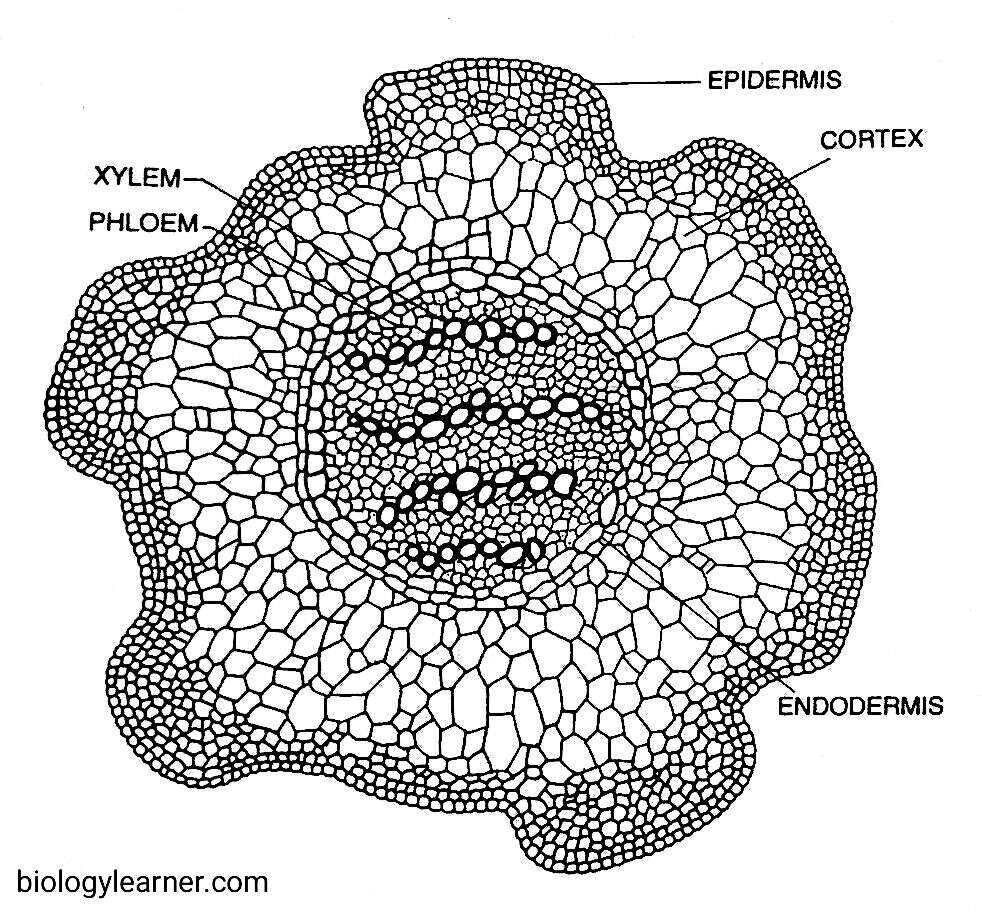
A single-layered endodermis is present just below the cortex. It has characteristic casparian strips on the radial walls. At the maturity of the stem, the endodermis may not be clearly recognisable as the endodermal cells become thickened.
The endodermis is followed by the pericycle, which is composed of 2–6 layers of compactly arranged thin-walled parechymatous cells.
Stele
The stele is protostelic (i.e., pith is absent) in nature and consists of only primary xylem and primary phloem. However, the arrangement of the vascular tissues, i.e., xylem and phloem, and associated parenchyma within the stele, varies greatly from species to species as well as in different portions of the same plant.
In L. phlegmaria, L. serratum, etc., the stele is an actinostele, in which the xylem is star-shaped. The protoxylem is situated at the periphery (exarch). The phloem is present in isolated masses, alternating with the xylem. In L. annotinum, the stele is also actinostelic, but the furrows in the xylem are much more and show a stellate arrangement.
In L. volubile, the stele is a plectostele. Here, the xylem, in a transverse section, appears to be in the form of separate plates, and the phloem not only surrounds the xylem but also is distributed between the plates of the xylem. The plectostele is also seen in L. densum and L. clavatum.
In L. laterale, L. drummondii, L. cernuum, etc., the xylem and the phloem are distributed uniformly. But the xylem is present in irregularly scattered groups, embedded in the ground mass of phloem (Ogura, 1938). This type of protostele is called the mixed protostele.
The xylem is exarch in all the species and consists only of tracheids. The tracheids are annular or spiral in the protoxylem but scalariform in the metaxylem.
The phloem is composed of sieve tubes and phloem parenchyma.
Anatomy of Root
A transverse section of the root of Lycopodium also shows an outermost epidermis, a cortex, and a stelar region, like that of the stem.
The epidermis is single-layered and consists of thin-walled parenchymatous cells. Many unicellular root hairs arise from the epidermal cells of the apical region of the root. These hairs are formed in pairs.
Below the epidermis is the broad cortex, which is several layers thick. It is composed of parenchymatous cells.
The outer few layers of cells often become heavily sclerified as the root gets older. These sclerified cells provide mechanical support to the root.
The center of the root is occupied by the stele. The stele is protostelic with monarch to polyarch xylem. It is generally monarch with one mass of protoxylem in young roots and becomes diarch or triarch in older roots.
In some prostrate species (e.g., L. clavatum), the stele is polyarch, i.e., having 6–10 strands of xylem arranged radially. The phloem lies between the radiating arms of the xylem.
In L. selago and L. pithyoides, the xylem is diarch. When the xylem is diarch, it usually appears as C, U, or crescent-shaped with protoxylem at the tips. The phloem is present between the two ends of the xylem as a single mass.
The stelar region is surrounded by endodermis and pericycle. The endodermis and pericycle may be indistinct at maturity.

Anatomically, the cortical roots are exactly similar to those of normal roots. In the cortical roots, however, the epidermis is poorly developed and the root hairs are absent.
Anatomy of Leaf
The leaf of Lycopodium is very simple in structure. In a cross-section, it shows an outermost epidermis, an undifferentiated mesophyll, and a single central vascular bundle.
The epidermis is one cell thick and consists of parenchymatous cells. These cells are highly cutinized on their outer walls.
The epidermis is interrupted by the stomata. In most species (e.g., L. segalo, L. clavatum), the stomata are present on both surfaces of the leaf, i.e., amphistomatic. However, in species with diamorphic leaves (e.g., L. volubile), the stomata are mostly confined to the lower surface, i.e., hypostomatic.
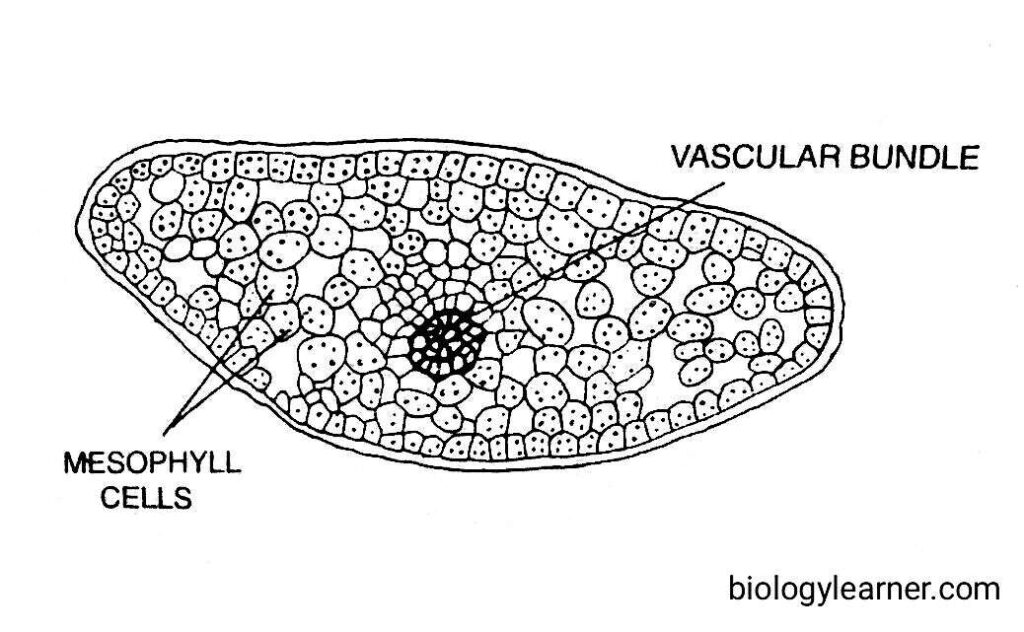
The mesophyll lies between the epidermis and the vascular bundle. It is usually undifferentiated with rounded or angular cells. The cells may have intracellular spaces.
In the centre of the leaf is a single concentric vascular bundle with xylem surrounded by phloem. The xylem is made up of thick-walled annular or spiral tracheids. Whereas phloem consists of phloem parenchyma and narrow sieve cells.
The vascular bundle remains surrounded by a sclerenchymatous sheath.
Reproduction in Lycopodium
The sporophyte of Lycopodium reproduces vegetatively as well as by the formation of spores.
Vegetative Reproduction
Vegetative reproduction is quite common in Lycopodium and occurs through the following methods:
Gemmae or Bulbils
The gemmae, or bulbils, are bud-like, modified lateral branches which usually develop in large numbers on the apex of the stem (e.g., L. phlegmaria, L. selago, and L. lucidulum).
The morphology of bulbils is still not fully known. Each bulbil consists of a short, reduced axis surrounded by many fleshy leaves. The leaves are rich in reserve food materials.
On falling to the ground, the bulbils grow into new sporophytic plants.
Fragmentation
Fragmentation can be seen in creeping species, e.g., L. incondatum.
In this method, the progressive death and decay of the older parts of the stem reach the region of branching, and the young branches get separated. Each such branch grows independently and develops into a new Lycopodium plant.
Resting Buds
In Lycopodium inundatum, the entire plant body except the growing tip of the rhizome or stem is dead during the unfavourable condition (i.e., winter). This tip of the rhizome or stem stores reserve food and functions as a resting bud.
With the onset of favourable conditions, the resting bud resumes growth and gives rise to a new individual.
Root Tubercles
In a few species, such as L. cernuum and L. ramulosum, bud-like small tubercles are developed in the apical region of the adventitious root by the proliferation of the parenchymatous cells (Holloway, 1917).
These tubercles are capable of germinating into new sporophytes.
