Polysiphonia is a filamentous marine red alga of the family Rhodomelaceae.
- Polysiphonia is mainly found along the coasts of the Atlantic and Pacific Oceans.
- The plant body of Polysiphonia remains attached to a rocky surface or other algae by rhizoids.
- It shows polysiphonous organisation.
| Class: | Rhodophyceae |
| Sub-Class: | Florideae |
| Order: | Ceramiales |
| Family: | Rhodomelaceae |
| Genus: | Polysiphonia |
Salient Features of Polysiphonia
The salient features of Polysiphonia are as follows:
- The plant body is red, profusely branched, filamemtous, and polysiphonous.
- The thallus is heterotrichous in nature and consists of two types of filaments: prostrate filaments and erect filaments.
- The prostrate filament creeps over the substratum by means of rhizoids or holdfasts.
- Branches are dimorphic, i.e., two types of branches can be seen in Polysiphonia: small branches and long branches.
- Small branches, or trichoblasts, bear sex organs.
- All cells are connected to each other by pit connections (cytoplasmic connections).
- Cells contain many pigments, such as chlorophyll-a, chlorophyll-d, β-carotenes, xanthophylls, phycocyanin-R, and phycoerythrin-R, in the chromatophores.
- Floridean starch is the reserve food material.
- Apical growth takes place by means of a single dome-shaped apical cell.
- Sexual reproduction is of the advanced oogamous type.
- The male sex organs are spermatangia, produced in dense clusters upon the fertile trichoblast of the male gametophyte.
- The female sex organs are called carpogonia or procarps. They are produced terminally upon a greatly reduced fertile trichoblast in the female gametophyte.
- The zygote shows an elaborate post-fertilization stage, forming cystocarp.
- Cystocarp contains carposporangia, which produce diploid carpospores.
- Presence of an isomorphic alternation of generations with a triphasic life cycle (i.e., a haploid gametophyte, a diploid carposporophyte, and a diploid tetrasporophyte).

Occurrence of Polysiphonia
- Polysiphonia is a genus of about 200 species.
- It is found exclusively in marine habitats and is widely distributed all over the world.
- Most of the species (e.g., P. elongata) prefer to grow on rocks as lithophytes.
- Some species are epiphytic on other plants and algae; e.g., P. ferulacea occurs on Galidium pusillum, P. urceolata grows on the thallus of Laminaria, and P. variegata is found on the roots of mangroves.
- Polysiphonia fastigiata occurs as a semiparasite on some brown algae, such as Fucus and Ascophyllum nodosum.
- A few species (e.g., P. variegata) grow in polluted brackish waters near the estuaries of the sea.
- Species of Polysiphonia also occur abundantly in shallow, stagnant water along the coasts of the Atlantic and Pacific Oceans.
Thallus Structure of Polysiphonia
The plant body of Polysiphonia is uniaxial (i.e., one main axis and all the others are side branches), branched, filamentous, and generally red or purple-red in colour. The filaments are profusely branched and consist of numerous siphon-like cells arranged in definite tiers. Hence, the name given to this genus is Polysiphonia.
Due to continuous branching and re-branching, the plant appears as a bush. The bush is generally small but may reach up to 30 cm in height.
In most species, the thallus exhibits a heterotrichous habit. It is differentiated into a prostrate system and an erect system.
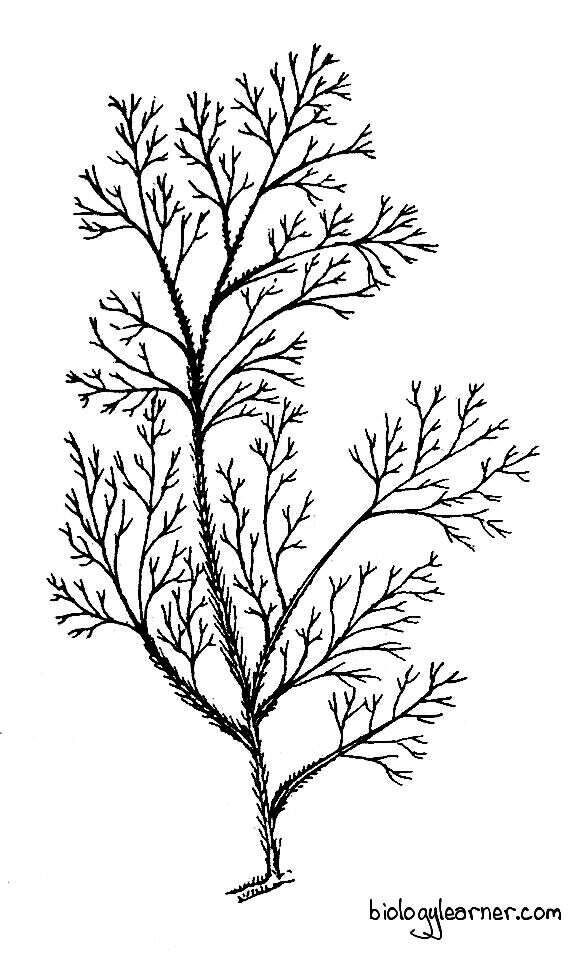
Prostrate System
- The prostrate system is basal and creeps over the substratum.
- In many Polysiphonia species (e.g., P. nigrescens, P. urceolata), it may be multiaxial and well developed.
- The prostrate system is attached to some suitable substratum by many rhizoids.
- The rhizoids are well developed, thick-walled, and unicellular. They are much lobed at the apex and form definite attachment discs.
- In a few species, such as P. violacea and P. elongata, the multiaxial prostrate system is absent, and the rhizoids develop from the lowermost cells of the erect system.

Erect System
- The erect filaments of the thallus develop from the prostrate system. The erect system is feathery in appearance and is made up of a main axis from which many branches are developed.
- The branches are of two types: long branches (branches of unlimited growth) and short branches (branches of limited growth).
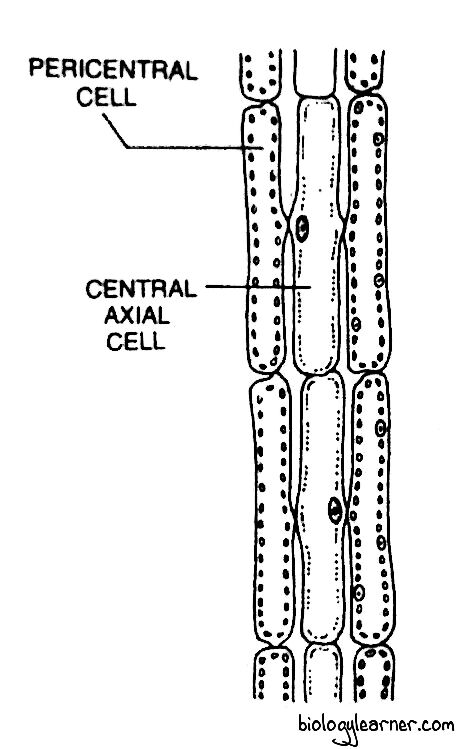
- The main axis and long branches consist of a central siphon (large central filament) of many elongated cylindrical cells situated in a vertical row.
- The central siphon is surrounded by 4–20 smaller pericentral cells, or pericentral siphons.
- The cells of the central siphons and pericentral siphons remain connected with one another through pit connections (cytoplasmic connections). Hence the complete plant body is polysiphonous in structure.
- In most of the species, the pericentral syphon is covered by 3-layers of cortical cells. They formed due to periclinal and anticlinal divisions of the cells of the pericentral siphon in the older branches.
- The cortical cells are also called cortical siphons and are parenchymatous in nature. They may be present in the lower part of the thallus (e.g., P. mollis) or throughout the thallus (e.g., P. crassiuscula).
- The branches of Polysiphonia are lateral and monopodial. The long branches (branches of unlimited growth) are called long lateral branches, while the short branches (branches of limited growth) are called trichoblasts.
Short Branches or Trichoblasts
The trichoblasts are uniaxial (monosiphonous) in structure and lack pericentral siphons. They are spirally arranged, dichotomously branched, and fine hair-like. Due to the absence of chromatophores, trichoblasts are colourless.
The trichoblasts arise on both the main axis and long branches. The short branches are deciduous. Hence, perennial species shed these branches before winter and develop again in the spring.
Some trichoblasts bear sex organs and are known as fertile trichoblasts.
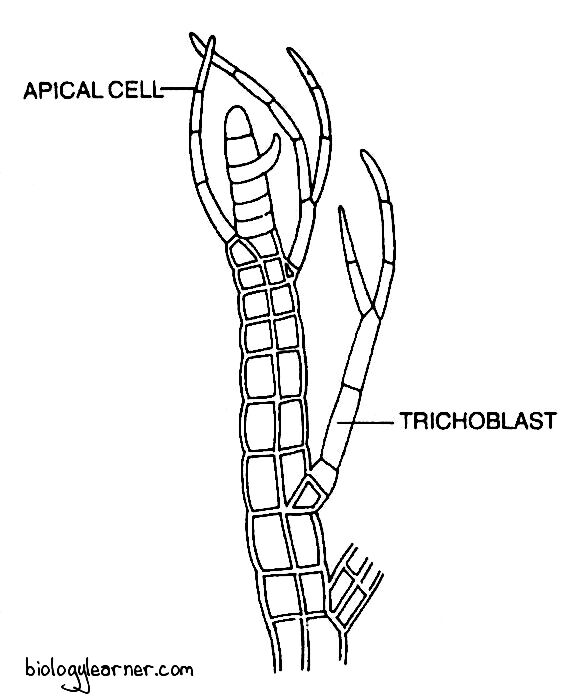
Development of Trichoblasts
- The trichoblast develops from a single trichoblast initial.
- The trichoblast initial is differentiated by the oblique division of the apical cell.
- It is a small cell and lies 2–3 cells below the apical cell.
- The small trichoblast initial divides repeatedly to form a dichotomously branched, multicellular, uniseriate, hair-like trichoblast.
Long Lateral Branches
The long lateral branches are similar to the main axis. They are polysiphonous (multiaxial) at the base and monosiphonous in the distal parts.
The long branches develop in radial or spiral symmetry.
Development of Long Branches
- The long branches arise from the basal cells of the short branches.
- In species like P. violacea, they occur as outgrowth from trichoblast initials.
- The long branches develop along with trichoblasts, and after a few divisions, the trichoblasts are pushed aside, so the branches appear to arise from trichoblasts dichotomously.
Cell Structure of Polysiphonia
- The cells of the central and pericentral siphons are elongated and cylindrical.
- Each cell is surrounded by a thick cell wall.
- The cell wall is differentiated into two layers: an outer pectic layer and an inner cellulosic layer.
- A thin plasma membrane encloses the cytoplasm. The cytoplasm contains a single nucleus and many discoid chromatophores.
- The chromatophores are devoid of pyrenoids and consist of pigments like chlorophyll-a, chlorophyll-d, β-carotenes, xanthophylls, phycocyanin-R, and phycoerythrin-R.
- Each cell possesses a large central vacuole in the cytoplasm. The vacuole is delimited by the membrane tonoplast.
- Floridean strach and floridoside are the reserve food materials found in the cytoplasm.
Growth of the Thallus
- Growth of Polysiphonia takes place by the dome-shaped apical cell, situated at the tip of the central siphon.
- The apical cell cuts off segments at its posterior surface by transverse divisions and forms the central siphon.
- Some of the lower cells then divide periclinally to form pericentral cells around the central siphon.
Reproduction in Polysiphonia
Polysiphonia shows an isomorphic triphasic alternation of generations. Both gametophytic (haploid) and sporophytic (diploid) generations are identical. Sexual reproduction takes place in the gametophytic plants, and asexual reproduction occurs in the sporophytic plants.
In the life cycle of Polysiphonia, three kinds of thalli are found. These are:
- haploid gametophyte
- diploid carposporophyte, and
- diploid tetrasporophyte
Gametophyte
- The gametophytic thalli are haploid, free-living, and dioecious, i.e., heterothallic (male and female sex organs are borne on different plants).
- The male sex organs (spermatangia) are produced on male gametophytic plants, while the female sex organs (carpogonia) are formed on female gametophytic plants.
Carposporophyte
- The carposporophyte is diploid and remains attached to the female gametophyte as a parasite.
- It develops from the zygote after fertilization.
- The carposporophyte reproduces asexually by the formation of diploid spores called carpospores.
Tetrasporophyte
- The tetrasporophytic plant is formed by the germination of diploid carpospores.
- It is diploid and independent.
- The tetrasporophyte reproduces asexually by producing haploid tetraspores.
- Tetrasporophytes and gametophytes (both male and female gametophytes) are morphologically similar but differ in producing the reproductive organs.
Sexual Reproduction
Sexual reproduction in Polysiphonia is of the advanced oogamous type. It occurs in the gametophytic thalli.
Most of the gametophytic thalli are heterothallic, i.e., dioecious. The sex organs are developed on different plants.
The male sex organs are spermatangia, borne on male plants. The female plants bear female sex organs called carpogonia or procarps.
Spermatangium
The spermatangium (antheridium) is the male sex organ in Polysiphonia. Many spermatangia are produced in dense clusters on the fertile trichoblasts near the apex of the male gametophytic plant.

- Initially, the male trichoblast develops as a side branch on the plant body.
- Each trichoblast, when it is only 2–3 celled, divides dichotomously and becomes branched.
- In most of the species, one branch remains sterile and the other bears spermatangia. In some species, both branches become fertile.
- The sterile branch may divide again to form fertile trichoblasts.
- The spermatangia bearing fertile trichoblasts are called spermatangial branches.
- During the development of the spermatangium, all cells of the uniaxial trichoblast except a few basal cells (i.e., 2–3 cells) divide periclinally and form pericentral cells on both sides.
- The pericentral cells may divide once or twice to form spermatangial mother cells on the outer side.
- Each spermatangial mother cell cuts off 2–4 spermatangia or antheridia. The complete structure forms a cone-shaped cluster of spermatangia.
- The mature spermatangium is a short-stalked spherical or oblong, unicellular structure.
- The cell wall of each spermatangium is differentiated into three layers: the inner reflective, the middle gelatinous, and the outer thick layer.
- The contents of the spermatangium metamorphose into a single, non-motile, male gamete, or spermatium.
- Each spermatium is liberated from the spermatangium by the formation of a narrow apical slit in the spermatangial wall.
- The liberated spermatia are dispersed by water currents.
- After the liberation of spermatia, new spermatangia may develop on the old spermatangia.

Carpogonium
The carpogonium (oogonium) is the female sex organ in Polysiphonia. It develops on a greatly reduced fertile trichoblast of the female gametophytic plant.

- During the development of the carpogonium, the female trichoblast initial arises from the central siphon, 2-4 cells below the apical cell.
- It divides repeatedly to form a reduced female trichoblast of 5–7 cells.
- The three lowermost cells of the female trichoblast divide vertically and form an ensheathing layer of pericentral cells, which are arranged in three tiers, one above the other.
- One of the pericentral cells in the middle tier on the adaxial side (that facing the main axis) becomes the supporting cell.
- The supporting cell cuts off a small initial cell at its free end (outside part), called the procarp initial.
- The procarp initial cell undergoes repeated divisions to form a small, curved, four-celled branch known as the carpogonial filament (carpogonial branch) or procarp.
- The terminal cell of the carpogonial filament functions as the carpogonial mother cell.
- The carpogonial mother cell further develops into the flask-shaped carpogonium.
- The carpogonium consists of a basal swollen portion, the carpogonial base in which the egg or female gamete is present and an upper elongated tubular portion called the trichogyne.
- Simultaneously, the supporting cell cuts off two sterile cells, one at the base, called the basal sterile filament initial, and another at the lateral side, the lateral sterile filament initial.
- The lateral sterile filament initial divides transversely to forms a 2-celled lateral sterile filament.
- The pericentral cells adjacent to the supporting cell start growing to cover the carpogonium (fertilised carpogonium).
- Later on, they form a sterile sheath (the protective covering) around the carpogonium, called the pericarp.
- The pericarp develops an opening at the tip, which is known as the ostiole.
Fertilization
- During fertilization, the spermatia are carried by the water currents to the carpogonium.
- As the spermatia reach the carpogonium, one of them adheres to the trichogyne.
- The walls between the trichogyne and the spermatium dissolve at the point of contact.
- The male nucleus enters the trichogyne and moves down to the swollen base of the carpogonium.
- Finally, the male nucleus fuses with the egg nucleus and forms the diploid zygote.
Post Fertilization Changes (Development of Cystocarp) in Polysiphonia
After fertilization, many changes occur simultaneously within and around the carpogonium of Polysiphonia. These changes lead to the development of the cystocarp.

- Immediately after fertilization, the 2-celled lateral sterile filament divides and becomes a short filament of 4–10 cells.
- The basal sterile initial divides to form a 2-celled basal sterile filament.
- Simultaneously, the supporting cell of the carpogonium divides transversely and develops a haploid auxiliary cell on its upper side, just below the basal portion of the carpogonium (carpogonial base).
- The auxiliary cell then establishes a tubular cytoplasmic connection with the carpogonial base.
- Now the diploid zygotic nucleus (2n) within the carpogonium divides mitotically into two diploid daughter nuclei.
- One of the nuclei migrates into the auxiliary cell through the tubular connection, and the other one remains in the carpogonium. Thus, the auxiliary cell contains one haploid nucleus (n) and one migrated diploid nucleus (2n).
- The haploid nucleus in the auxiliary cell now degenerates.
- The trichogyne gradually begins to shrivel.
- The diploid nucleus present in the auxiliary cell then divides mitotically and forms two daughter nuclei.
- One of the nuclei migrates into a small lateral outgrowth developed on the upper side of the auxillary cell. This small outgrowth on the auxiliary cell is called the gonimoblast initial.
- In this way, many gonimoblast initials can develop on the auxiliary cell.
- Now each gonimoblast initial divides by repeated mitotic divisions to form a short multicellular filament called the gonimoblast filament or gonimalobe.
- The terminal cell of the gonimoblast filament becomes swollen and develops into a pear-shaped carposporangium.
- The contents of the carposporangium metamorphoses into a single non-motile diploid carpospore.
- Meanwhile, the carpogonium, auxiliary cell, and supporting cell fuse together and form a single large, irregularly shaped nutritive cell called the placental cell, or central cell.
- The placental cell provides nourishment to the growing carposporophyte (i.e., carposporangia and carpospores).
- The pericentral cells of the female trichoblast adjacent to the supporting cell produce several vegetative filaments.
- Many gonimoblast filaments attached to the placental cell are covered by sterile vegetative filaments and develop an urn-shaped structure called the cystocarp.
- The outer covering of the cystocarp is a two-layered wall and is known as the pericarp.
- The pericarp is haploid and has an opening at the apex called the ostiole.
- The diploid part of the cystocarp represents the carposporophyte (carposporangia bearing carpospores). Thus, the cystocarp is a partly haploid and partly diploid structure.


Asexual Reproduction
Asexual reproduction in Polysiphonia takes place in the sporophytic plants. It occurs through the formation of carpospores and tetraspores.
Carpospores develop in the carposporophyte, while tetraspores produce in the tetrasporophyte.
Carpospores
- The carpospore is diploid, non-motile, uninucleate, and developed in the carposporophyte.
- The mature carposopres are liberated from the cystocarp through the ostiole and are carried away by water currents.
- On coming into contact with the suitable substratum, the carpospore secretes a wall around it and attaches itself to the substratum.
- It then divides by mitotic division to form a small lower cell and a large upper cell.
- Both of these cells again divide mitotically and form a four-celled small filament.
- The lowermost cell of the filament is called the rhizoidal cell.
- The rhizoidal cell elongates and forms the colourless rhizoids.
- The uppermost cell is dome-shaped and functions as the apical cell.
- The apical cell divides by transverse division and forms the central siphon cell.
- The central cell undergoes further division (periclinal division) to form pericentral cells and ultimately develop the polysiphonous body.
In this way, the carpospore germinates into the diploid tetrasporophytic plant.
Tetraspores
- The tetrasporophyte reproduces asexually by means of tetraspores.
- The tetraspores develop in the tetrasporangium.
- The tetrasporangium is a sac-like diploid structure.
- It is produced from the pericentral cell of the thallus near the apical region.
- The single pericentral cell of each tier, toward the apical region, functions as the tetrasporangial initial.
- This pericentral cell (tetrasporangial initial) is smaller than the other pericentral cells in the same tier.
- The tetrasporangial initial divides vertically and forms an outer cell and an inner cell.
- The outer cell further divides to form two or more cover cells.
- The inner cell functions as the sporangial mother cell.
- The sporangial mother cell then divides transversely into a lower stalk cell and an upper tetrasporangial cell.
- The tetrasporangial cell enlarges and develops into the tetrasporangium.
- The fertile branches bearing tetrasporangia become twisted and swollen.
- The diploid nucleus of the tetrasporangium undergoes meiosis, forming four haploid tetraspores, or meiospores.
- The spores are arranged tetrahedrally in the tetrasporangium.
- At maturity, the tetraspores are liberated by rupturing of the sporangial wall along with lifting of the two cover cells.
- The liberated tetraspores germinate into new haploid gametophytic plants.
- Due to the hetarothallic nature of Polysiphonia, two of the four tetraspores develop male plants, and the other two produce female plants.
