Chlamydomonas is a unicellular, motile, green alga of the class Chlorophyceae.
- It is widely distributed and found mostly in stagnant water.
- Chlamydomonas is one of the most primitive eukaryotes in the plant kingdom.
| Class: | Chlorophyceae |
| Order: | Volvocales |
| Sub-order: | Chlamydomonadineae |
| Family: | Chlamydomonadaceae |
| Genus: | Chlamydomonas |
Salient Features of Chlamydomonas
The salient features of Chlamydomonas are as follows:
- The plant body is biflagellate, unicellular, elliptical, spherical, or pear-shaped.
- The flagella are equal in length and are present at the anterior end of the thallus.
- Prominent cub-shaped chloroplast is present, with one large pyrenoid containing a starch sheath.
- Two contractile vacuoles and one orange-red eyespot are found at the base of each flagellum.
- Volutin grains are the main reserve food products.
- There is a palmella-stage present.
- Asexual reproduction takes place by biflagellate zoospores.
- Sexual reproduction takes place through isogamy, anisogamy, and oogamy.
Occurrence of Chlamydomonas
Chlamydomonas is a large genus that contains about 400 species. Most of the species are found in freshwater lakes, ponds, ditches, etc.
Terrestrial species grow on damp, moist soil. In alpine and arctic regions, Chlamydomonas nivalis grows on snow and provides the red coloring to the substratum, resulting in the red snow. The snow becomes red due to the presence of the red pigment haematochrome in C. nivalis.
Chlamydomonas yellowstonensis is found in the Yellowstone National Park in America and imparts a yellow-green color to snow.
Thallus Structure of Chlamydomonas
The plant body of Chlamydomonas is unicellular, biflagellate, motile, and generally green in color.
A single cell represents the thallus. Each cell is spherical, ellipsoidal, oval, oblong, or pyriform in shape. The size of the cells varies from 20 to 30 μm in length.
The majority of species have a wider posterior side and a pointed anterior side, which subsequently ends in an apical papilla.
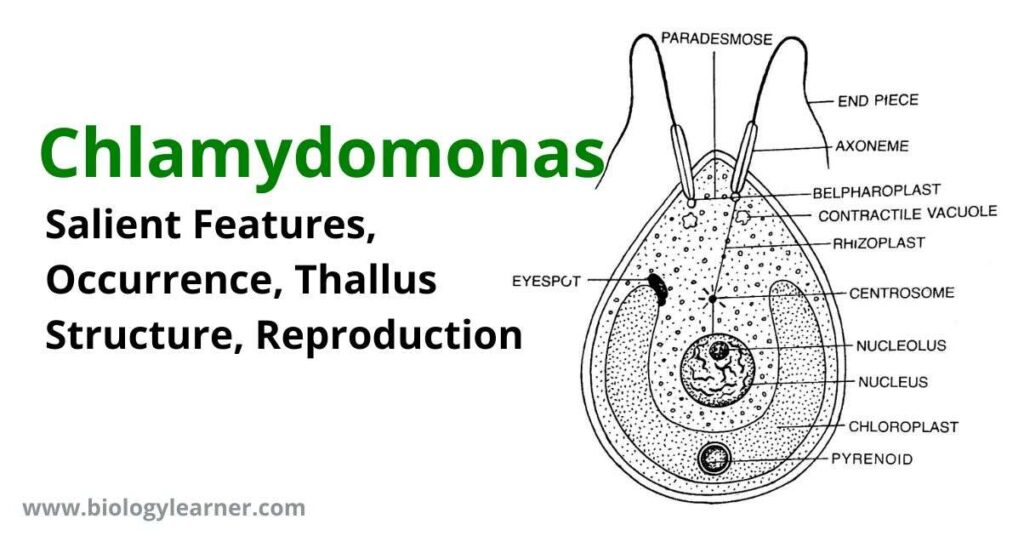
Under the microscope, the structure of the thallus shows the following parts:
- Cell Wall
- Cytoplasm
- Chloroplast
- Flagella
- Neuro-motor apparatus
- Stigma or Eyespot
Cell Wall
The cell is surrounded by a definite, smooth, and thin cell wall. It is composed of cellulose.
In some species (C. gleocystiformis), the cell wall is surrounded by a gelatinous pectin layer, which is formed by the dissolution of the pectose layer into pectin.
The gelatinous pectin layer protrudes at the anterior end of the cell to form a beak-like apical papilla.
The detailed structure of the cell wall shows that it is multilayered and made of cellulose fibrils. A plasma membrane is present inside the cell wall. It is semipermeable and consists of two membranes connected by an opaque zone. The cell membrane surrounds the cytoplasm.
Cytoplasm
The cytoplasm is present inside the plasma membrane of the thallus. It contains many cellular organelles such as the nucleus, mitochondria, endoplasmic reticulum, dictyosomes, ribosomes, etc.
A single large, dark nucleus remains suspended in the colorless part of the cytoplasm inside the cavity of the cup-shaped chloroplast.
The dictyosomes, or Golgi bodies, are present close to the nucleus.
The ribosomes in the cytoplasm are the 80S, whereas the 70S ribosomes characteristic of prokaryotic cells are found in the chloroplast.
Chloroplast
In Chlamydomonas generally, a single large, cup-shaped, or bowl-shaped parietal chloroplast is present in the cytoplasm. The chloroplast may be massive and occupy most of the protoplast.
However, in different Chlamydomonas species, the chloroplasts can be of various shapes. The chloroplast is H-shaped in C. bicilliata, parietal in C. mucicola, reticulate in C. reticulata, axile in C. steinii, and stellate in C. arachne.
The posterior end of the chloroplast generally contains a single pyrenoid covered with a starch sheath. But some species contain more than one pyrenoid. There are two pyrenoids in C. debaryana and many in C. gigantae.

The chloroplast has a double membrane envelope with lamellae arranged in the form of stacks. The stacks form the grana, and the matrix constitutes the stroma. DNA, ribosomes, and microtubules are present in the chloroplast matrix.
Stigma or Eyespot
The anterior side of the chloroplast on one side contains a tiny spot of orange or reddish color called the stigma or eyespot. It is a photoreceptive organ with a round to oval form.
The stigma consists of a curved pigmented plate, the pigmentosa, and a biconvex lens. Two or three parallel rows of fat droplets called granules containing carotenoids can be found on the plate. The lens is as thick as the cell wall and is photosensitive in function.
The stigma is sensitive to light and is supposed to be concerned with directing the movement of the flagella. If a beam of intense light is made to fall on the stigmatic surface, it is observed that the Chlamydomonas swim away from the light.
Flagella
There are two whiplash types of flagella present in the anterior portion of the thallus. They are equal in length. The flagella are usually longer than the thallus, but in some species, they can be equal to or shorter than the thallus.
Each flagellum originates from a basal granule or blepharoplast that is situated towards the anterior papillate or non-papillate region of the cytoplasm. The flagella emerge through very fine canals on the outer wall of the thallus.
Under an electron microscope, flagellum shows eleven components of fibrils with a typical 9+2 arrangement. The two central fibrils are single fibrils, while the nine peripheral fibrils are double fibrils.
Neuromotor Apparatus
In some species of Chlamydomonas (e.g., C. nasuta), a sensitive neuromotor apparatus is present.
The neuromotor apparatus is made up of two basal granules, or blepharoplasts, from which the flagella originate. The two blepharoplasts are joined by thin protoplasmic fibrils known as “Paradesmos“. One of the blepharoplasts is connected to the centrosome of the nucleus by a thread called “Rhizoplast”. Many fine fibrils interconnect the centrosome with the nucleolus.
The nucleus controls the flagellar movement through the rhizoplast in response to light, chemicals, and other stimuli.
Contractile Vacuoles
In Chlamydomonas, usually, two contractile vacuoles are present near the blepharoplast. The number and position of these vacuoles may vary in different species.
The contractile vacuoles can undergo alternate expansion and contraction. They function as excretory organs, controlling the water content of the cell by releasing more water at specific times. The osmotic regulation of the cell is also controlled by contractile vacuoles.
Reproduction in Chlamydomonas
Chlamydomonas reproduce by both asexual and sexual methods. Asexual reproduction occurs during favorable as well as unfavorable conditions. Sexual reproduction takes place only under favorable conditions.
Asexual Reproduction
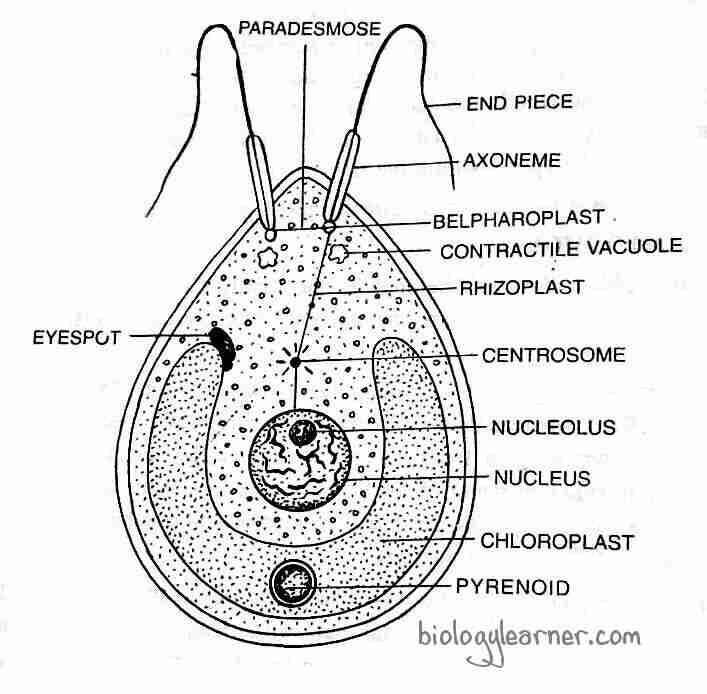
The asexual reproduction in Chlamydomonas takes place commonly by the Zoospore formation. Others, however, reproduce through aplanospores, hypnospores, and the palmella stage.
By Zoospores
During favorable conditions, zoospores are formed. At the beginning of the zoospore formation, the protoplast of the parent cell contrasts and gets separated from the cell wall. The parent cell discards the flagella, or in some species of Chlamydomonas, the flagella are absorbed and become non-motile. The contractile vacuoles and the neuro-motor apparatus also disappear at this stage.

The protoplast is divided repeatedly by simple mitotic divisions, producing 2-8 or more daughter protoplasts. Each unit of protoplast with a single nucleus secretes a wall around it and develops contractile vacuoles and flagella at the anterior end. These flagellated daughter protoplasts are called zoospores or swarmer.
The zoospores are identical to the parent cells in structure but smaller in size. The motile zoospores are liberated from the parent cell by dissolution or gelatinization of the parent cell wall and form new Chlamydomonas.
By Aplanospores
The aplanospores are formed during unfavorable conditions. In some species (C. caudata), the parent cell loses flagella and becomes non-motile, like in zoospore formation.
The protoplast divides repeatedly and forms 2 to 16 daughter protoplasts. The daughter protoplasts become rounded and secrete a thin wall around them. These non-motile, thin-walled structures are called aplanospores.
During favorable conditions, aplanospores may germinate either directly or divide to produce zoospores.
By Hypnospores
During extremely unfavorable conditions such as severe drought, the aplanospore-like structures in some Chlamydomonas species (e.g., C. nivalis) are enveloped by a thicker wall with abundant food reserves, and the structures are called hypnospores.
Each hypnospore germinates after a resting period to give rise to a new plant or divides to produce zoospores.
Palmella Stage
If the environmental conditions become unfavorable, such as shortage of water, excess of salts, etc., the zoospores alter their formation and are not able to get out of the parent cell. They remain involved in a gelatinous sheath. The daughter protoplasts form a gelatinous wall around themselves without developing flagella.
The non-motile daughter cells may divide again and form an amorphous colony. This amorphous colony looks like another green alga, Palmella of Tetrasporales, and is called the Palmella stage.
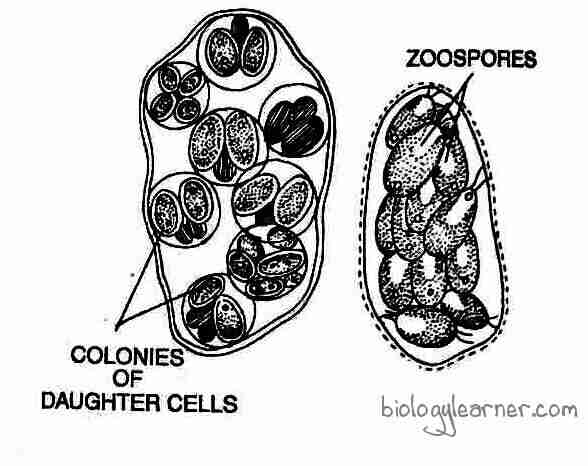
With the onset of favorable conditions, the gelatinous wall is dissolved and the cells develop flagella, becoming motile. Each motile daughter cell escapes from the gelatinous sheath and later develops into a new plant.
The palmella stage is commonly observed in C. kleinii and C. braunii.
Sexual Reproduction
In Chlamydomonas, sexual reproduction is highly variable and can be isogamous, anisogamous, or oogamous. Chlamydomonas may be monoecious (homothallic), i.e., both types of gametes (male and female) are produced in the same thallus (e.g., C. mogama and C. media) or maybe dioecious (heterothallic), i.e., male and female gametes come from the distinct plants (e.g., C. reihardii, C. moewusii).
Nitrogen content desiccation or deficiency during sexual reproduction favors gamete formation. Gamete formation takes place by the repeated divisions of the protoplast in the same way as in zoospore formation.
In the majority of species, the gametes are very small, unicellular, uninucleate, and biflagellate structures. The gametes may be naked without a cell wall and are called gynogametes (e.g., C. debaryana). When the gametes are covered by a cell wall, they are called calyptogametes (e.g., C. media).
Isogamy
The majority of the Chlamydomonas species (e.g., C. media, C. monoica, C. debaryana, etc.) are isogamous. In this type, the fusion takes place between the morphologically identical gametes. The gametes are similar in size, shape, and structure. They are morphologically similar but physiologically dissimilar.
Generally, the vegetative cell divides into 8 to 64 gametes. In some isogamous species (e.g., C. snowiae), the vegetative cell may directly function as a gamete without undergoing any division. this type of fusion is called hologamy (Smith, 1955).
In C. snowiae, the thalli lose their walls and function as gametes. The two gametes come close to each other by their anterior ends, and later fusion occurs on their lateral sides. After the fusion of two gametes, a quadri-flagellate and bi-nucleate structure i.e., the zygote is formed with two pyrenoids and two eyespots. The quadri-flagellate zygote remains motile for several hours to a few days.
In C. eugametos, the vegetative cells do not lose their walls; after fusion, the contents of one gamete enter another gamete.
According to Chapman (1964), isogamous reproduction occurs by the production of 8, 16, or 32 bi-flagellated gametes.
In this process, the vegetative thallus functioning as a gametangium comes to rest and loses its flagella. The protoplast gets separated from the cell wall.
The protoplast is divided repeatedly by longitudinal mitotic divisions, producing 8-16-32 or 64 daughter protoplasts. Each daughter protoplast forms a pair of flagella and transforms into a gamete. The gametes are liberated from the gametangium by breaking the gametangium wall.
The flagella of gametes are enclosed by agglutins (not found on the flagella of the vegetative cell) and secrete a hormone called gamone. These chemical substances help in the recognition of gametes of the opposite strains.
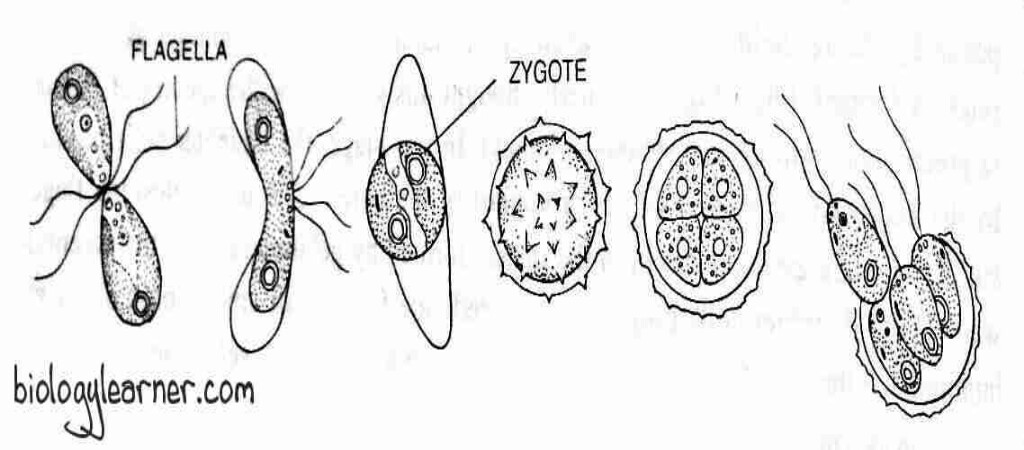
In heterothallic species (+) and (-) strains, gametes cluster together, and this phenomenon is known as clumping. The gametes of the opposite strain fuse at the anterior end (called apical fusion), or laterally (called lateral fusion). The paired gametes move away from the clump.
The wall at the point of contact dissolves, and fertilization takes place in two steps: plasmogamy and karyogamy. The fusion of cytoplasm occurs in plasmogamy, whereas the fusion of nuclei occurs in karyogamy.
After fertilization, a diploid quadriflagellate zygote is developed. Later on, the zygote loses its flagella and is enclosed in a thick wall known as a zygospore.
Anisogamy
Anisogamous sexual reproduction is observed in C. braunii, C. suboogama, etc.
In anisogamy, the fusing gametes (male and female) are unequal in size. The gametes are formed in different gametangia. Mostly, larger gametes are less active and formed in smaller numbers, while smaller gametes are more active and formed in relatively large numbers.
The smaller gametes are called male gametes or microgametes, whereas the larger gametes are called female gametes or macrogametes. Microgametes are developed in the male gametangium, in which the protoplast divides to produce 8 to16 gametes. Macrogametes are formed in the female gametangium, where the protoplast divides to produce 2 to 4 gametes.
The more active microgamete comes close to the macrogamete. The protoplast of the microgamete enters the macrogamete and fusion takes place. After fusion (plasmogamy and karyogamy), a diploid zygote (2n) is formed. Later, the zygote secretes a thick wall and transforms into a zygospore.
Oogamy
Oogamous sexual reproduction occurs in more advanced Chlamydomonas species, e.g., C. coccifera and C. ooganum.
The vegetative thallus (female cell) enlarges in size, becomes rounded, withdraws its flagella, and directly functions as a non-motile female gamete, macrogamete, or egg. Another cell functions as a male cell, which undergoes four divisions and forms 16 male gametes, or microgametes. The male gamete is quite distinct from the female gamete. It is motile, biflagellate, very small in size, and spherical in shape.
The active male gamete comes very close to the non-motile female gamete and attaches to the anterior end. The contact wall at the point of union dissolves and fertilization takes place. After fertilization (plasmogamy and karyogamy), a diploid zygote is formed.

Later, the zygote secretes a thick wall and transforms into a zygospore.
Zygote or Zygospore
The zygote initially remains motile for several hours (e.g., C. paradoxa) to about 15 days (e.g., C. pertusa). After losing flagella, it comes to rest on the substratum (actually, the zygote is a resting diploid spore) and secretes a multilayered thick wall.
As the zygote matures, it accumulates a large amount of oil and starch. Due to the presence of haematochrome, the zygospores are red in color. The zygospore can survive for long periods of time in unfavorable conditions.
Germination of Zygote
With the onset of favorable conditions, at the end of the resting period, the zygospore begins to germinate. During germination, the diploid zygote nucleus (2n) undergoes meiosis to form 4 haploid nuclei (n).
The four haploid nuclei with protoplasts make four haploid zoospores or meiozoospores. The segregation of sex (+ and – strains) takes place during meiosis. In four meiozoospores, two may be of the (+) type and two of the (-) type in heterothallic forms.
In some Chlamydomonas species, 8 or more meiozoospores are formed. The number of meiozoospores in a zygospore is 8 in C. reinhardtii and 16 to 32 in C. intermedia.
The inner zygote wall dissolves, and the haploid meiozoospores are liberated by breaking the outer wall. During the liberation, each meiozoospore contains an eyespot, a neuromotor apparatus, two flagella, and contractile vacuoles. The meiozoospores grow into new Chlamydomonas plants.
Life Cycle
Chlamydomonas, like other chlorophycean algae, has a haplontic life cycle with the zygote as the only diploid structure. The haploid Chlamydomonas thallus is unicellular, biflagellate, elliptical, spherical, or pear-shaped.
Chlamydomonas reproduce both asexually and sexually.
Asexual reproduction takes place, usually by zoospore formation. Others, however, reproduce asexually through aplanospores, hypnospores, and the palmella stage.
Sexual reproduction is highly variable and can be isogamous, anisogamous, or oogamous in type. Chlamydomonas can be monoecious or dioecious.
A diploid quadriflagellate zygote is formed after the fusion of male and female gametes.
The zygote goes through a period of rest before germination. During germination, the zygote undergoes reduction division, resulting in the formation of a haploid meiozoospore.
The meiozoospore grows into a new Chlamydomonas thallus.
