Ascobolus is a coprophilous fungus (i.e., it grows on animal dung) of the class Discomycetes.
The genus Ascobolus was introduced in 1791 by C.H. Persoon.
Occurrence of Ascobolus
Ascobolus is a genus of about 60 species. It occurs all over the world.
Most of the species are corpophilous, growing on the dung of various herbivorous animals. A few species also grow on other places, such as burnt forest soil, decaying plant materials, etc. Ascobolus carbonarius occurs on burnt soil.
- Some Ascobolus species are: A. indicus, A. carbonarius, A. viridis, A. immersus, A. magnificus, etc.

Read also- Rhizopus: Occurrence, Structure, Reproduction
Vegetative Structure of Ascobolus
The vegetative plant body of Ascobolus is a well-developed, perennial mycelium. The mycelium is composed of profusely branched septate hyphae.
The hyphae ramify within the substratum, forming a cottony appearance.
The hyphal cells are short and multinucleate. The microfibrillar cell wall surrounds each cell. It consists mainly of chitin.
The cytoplasm contains many nuclei, mitochondria, ribosomes, endoplasmic reticulum, oil droplets, and other substances.
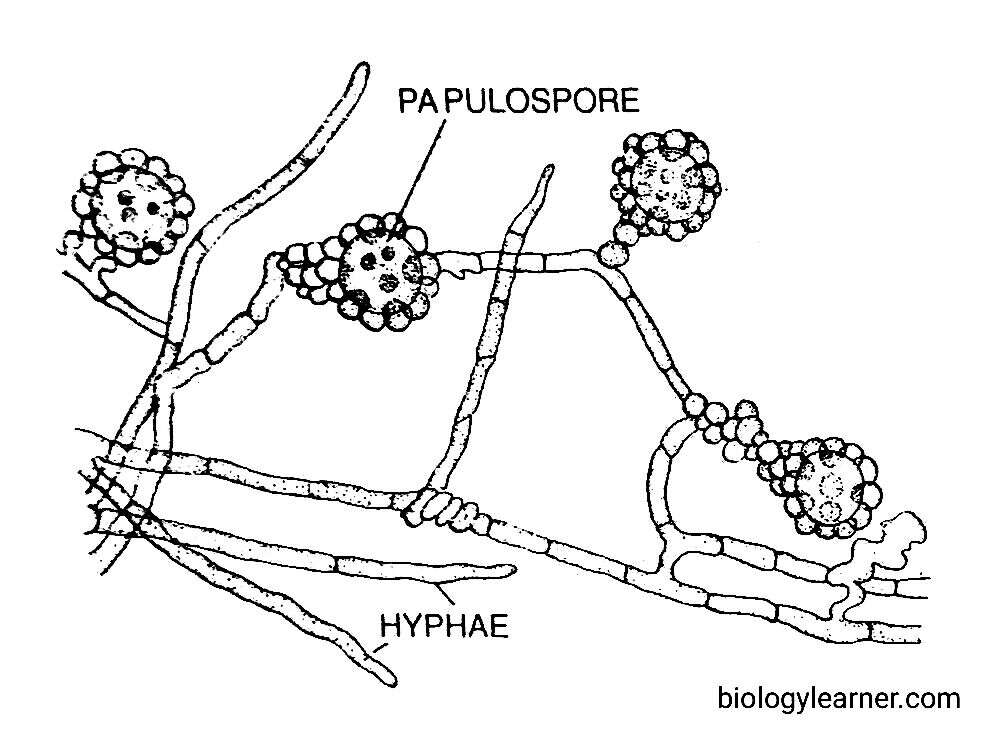
Reproduction in Ascobolus
Ascobolus reproduces by both asexual and sexual methods.
Asexual Reproduction
Asexual reproduction is rare in Ascobolus. It takes place by the formation of oidia (i.e., arthospores) and papulospores.
Some species, like A. denudatus and A. furfuraceus, produce oidia in chains on small hyphal branches (Webster, 1980). The oidia are thin-walled structures (hyphal cells). They can germinate to form new mycelia. It is reported that the oidia play an active role in sexual reproduction.
In A. scatigenus, papulospores are formed (Dodge, 1920). These are sclerotium-like structures, having one or two central cells surrounded by a covering hypha. This covering hypha later forms blister-like outgrowths. On germination, each papulospore may develop into a new hypha.
Sexual Reproduction
Sexual reproduction in Ascobolus is of the oogamous type. The male sex organs are called antheridia, while the female sex organs are known as ascogonia.
Ascobolus may be homothallic (e.g., A. crenulatus) or heterothallic (e.g., A. immersas, A. stercovarius, and A. furfuraceus).
A. scatigenus is a monoecious species (i.e., homothallic), capable of bearing both male and female sex organs but functionally heterothallic. This species shows two complementary strains (mycelia). Both strains develop antheridia and ascogonia. But the antheridia of one strain do not fertilise the ascogonia of the same strain. Hence, each strain is self-incompatible.
When the two complementary strains are intermingled, the sexual branches may be scattered singly or more often associated in pairs of 3 to 4. The paired branches elongate, one of which elongates more than the other. The shorter one is the male or antheridial branch, while the more elongated one is the female or ascogonial branch. Soon after, both antheridial and ascogonial branches become septate.
The antheridial branch is cylindrical or clavate and remains erect. Later on, it cut off an apical antheridium.
The ascogonial branch gradually coils the antheridium. It becomes more septate and is differentiated into a terminal long septate multicellular trichogyne, a large globose unicellular multinucleate ascogonium, and a multicellular stalk.
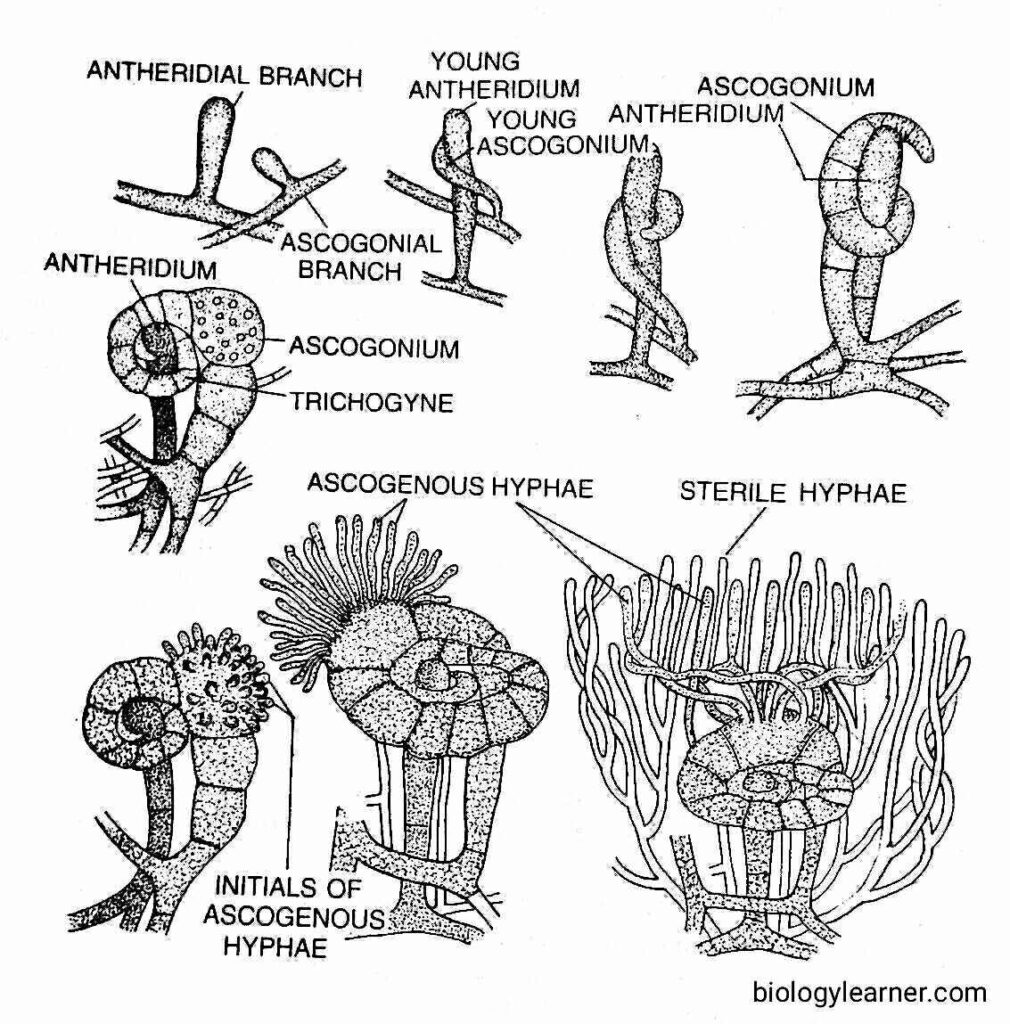
The trichogyne elongates rapidly and coils twice or thrice around the antheridium. The apical point of the trichogyne attaches to the tip of the antheridium.
The common wall dissolves at the point of their contact (plasmogamy), and the male nuclei pass to the ascogonium through the trichogyne.
The male and female nuclei form pairs towards the periphery of the ascogonium (Dodge, 1920). Soon, the ascogenous hyphae are developed from the fertilized ascogonium.
Simultaneously with the development of ascogenous hyphae, some sterile hyphal branches develop from the stalks of both the sex organs and also from the neighbouring mycelium. These branches subsequently intermingle with the ascogenous hyphae and ultimately form a cup-shaped fruit body called the apothecium (i.e., ascocarp).
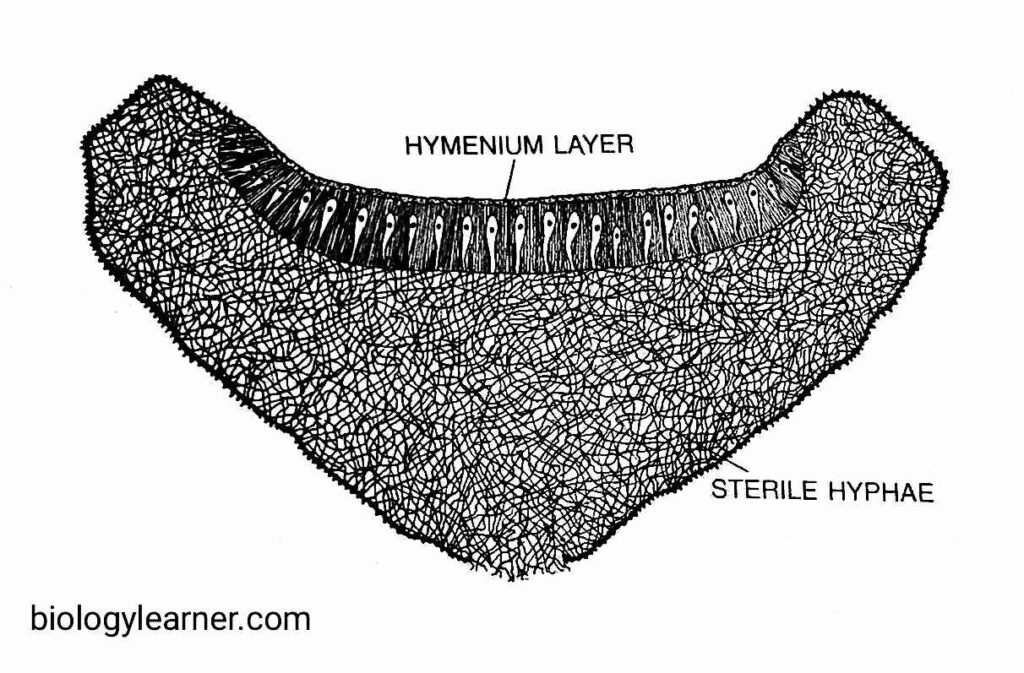
Apothecium
The apothecium is yellowish in colour. It is usually small, about 5 mm in diameter, but more than 2.5 cm in A. scatigenous.
The ascogenous hyphae increase in size, and a pair of nuclei migrate inside each of them. The transverse walls appear, and the hyphae become septate.
The apical cell of ascogenous hyphae divides to form four nuclei. These four nuclei are arranged in three-celled structures, i.e., a terminal uninucleate hook cell, a binucleate penultimate cell, and a uninucleate stalk cell (crozier formation). The two nuclei of the penultimate cell fuse (karyogamy), and thus a young ascus is formed.
In the young ascus, the diploid nucleus divides first by meiosis and then mitosis to form eight haploid nuclei. Each of the nuclei transformed into an ascospore.
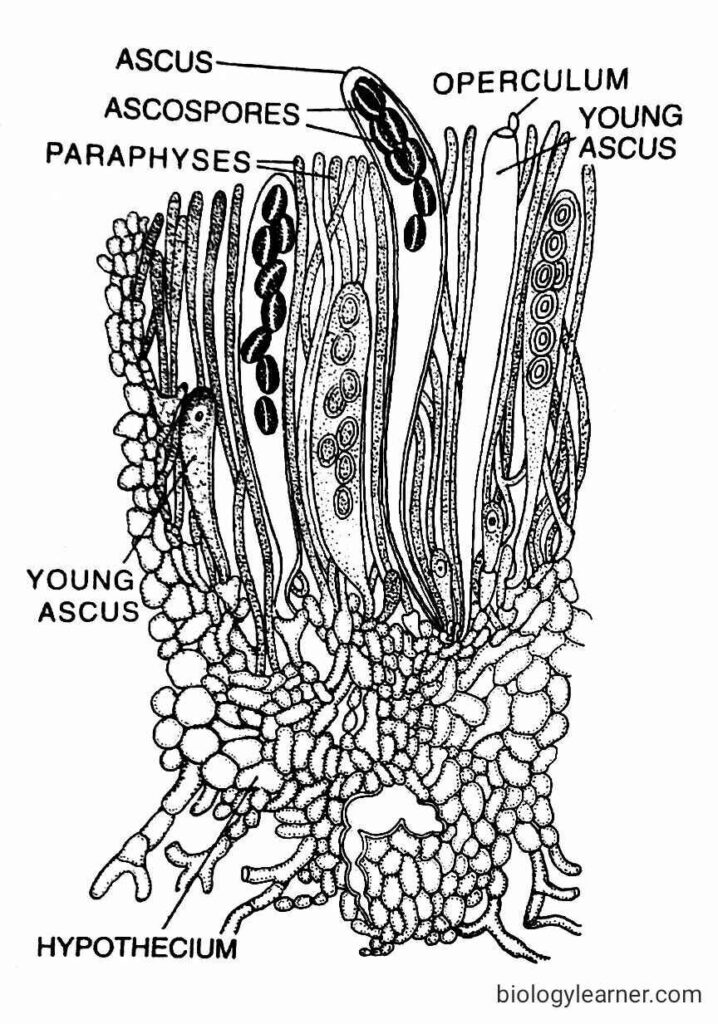
Each mature ascus is long and clavate, with eight ascospores, of which four belong to one strain and the other four belong to the opposite stream. There is an operculum, or lid, present at the apical end of the ascus.
The asci remain surrounded by septate paraphyses in the hymenium (upper region) of the apothecium. As the asci mature, the surface of the apothecium is studded with purple dots.
In Ascobolus, the development of apothecium is of two types (Van Brummelen, 1967).
- Cleistohymenial or Angiocarpic Type: In this type, the hymenium is enclosed in its early stage of development, e.g., A. furfuraceous.
- Gymnohymenial or Gymnocarpic Type: Here, the hymenium remains exposed from the very beginning, e.g., A. scatigenus.
Ascospores
The ascospores are haploid, uninucleate, and elliptical in shape. The size of the ascospores varies from 14–25 μm × 7–11 μm. But in A. immersus, they are very large, measuring 70 μm × 30 μm.
Each ascospore remains surrounded by two wall layers. The outer wall is ornamented and purple in colour, which later turns brown to black.
Liberation of Ascospores
The ascospores are liberated from the ascus through the operculum. On germination, each ascospore develops into new mycelium.
Taxonomic Position of Ascobolus
| Division: | Eumycota |
| Sub-Division: | Ascomycotina |
| Class: | Discomycetes |
| Order: | Pezizales |
| Sub-Order: | Pezizineae |
| Family: | Ascobolaceae |
| Genus: | Ascobolus |
