Volvox is a free-floating freshwater planktonic green alga of the class Chlorophyceae.
- It is usually found in stagnant water, such as ponds, pools, etc.
- Volvox diverged from its unicellular predecessors approximately 200 million years ago.
| Class: | Chlorophyceae |
| Order: | Volvocales |
| Sub-order: | Chlamydomonadineae |
| Family: | Volvocaceae |
| Genus: | Volvox |

Salient Features of Volvox
The salient features of Volvox are as follows:
- The plant body is a multicellular motile coenobium and has a globose or spherical hollow structure.
- The coenobium is composed of a large number of biflagellate, pear-shaped cells.
- The cells in a coenobium are interconnected by protoplasmic strands and are arranged in a single layer towards the periphery.
- Young coenobium contains only vegetative cells, which are primarily concerned with food production and locomotion.
- Mature coenobium contains vegetative cells as well as reproductive cells. Reproductive cells differ from vegetative cells.
- Starch is the reserve food material.
- Asexual reproduction occurs through specialized, enlarged cells called gonidia.
- Sexual reproduction is of the oogamous type, and the coenobia may be homothallic (e.g., V. globator) or heterothallic (e.g., V. aureus).
- The female gametes are large and non-motile, produced singly within the oogonium.
- The male gametes are spindle-shaped, narrow with a pair of apical cilia, and are produced in bunches within the antheridium.
Occurrence of Volvox
Volvox is a genus of about 20 species, which occur in colonies. It grows as plankton on the surface of water bodies like temporary and permanent freshwater tanks, ponds, pools, and lakes.
The Volvox colonies appear as minute floating balls on the surface of the water. During the spring and rainy seasons, the water surface becomes green due to its rapid growth.
Thallus Structure of Volvox
The plant body of Volvox is a motile coenobium (a colony with a definite shape and number of cells). The coenobium is a hollow sphere of mucilaginous substance.
At the periphery of the hollow sphere, a specific number of cells are arranged in a single layer. The number of cells may vary from 500 to 60,000 or more in different species (500-1,000 in V. aureus, 1500-20,000 in V. globator).
A Volvox cell is typical of the Chlamydomonas type in structure (except for a few like V. rouseletti and V. globator, which are of the Sphaerella type). Each cell is biflagellate and spherical, elliptical, or oval in shape, with a narrow anterior end and a broad posterior end. The two flagella are equal, whiplash-type, and are attached to the anterior end.
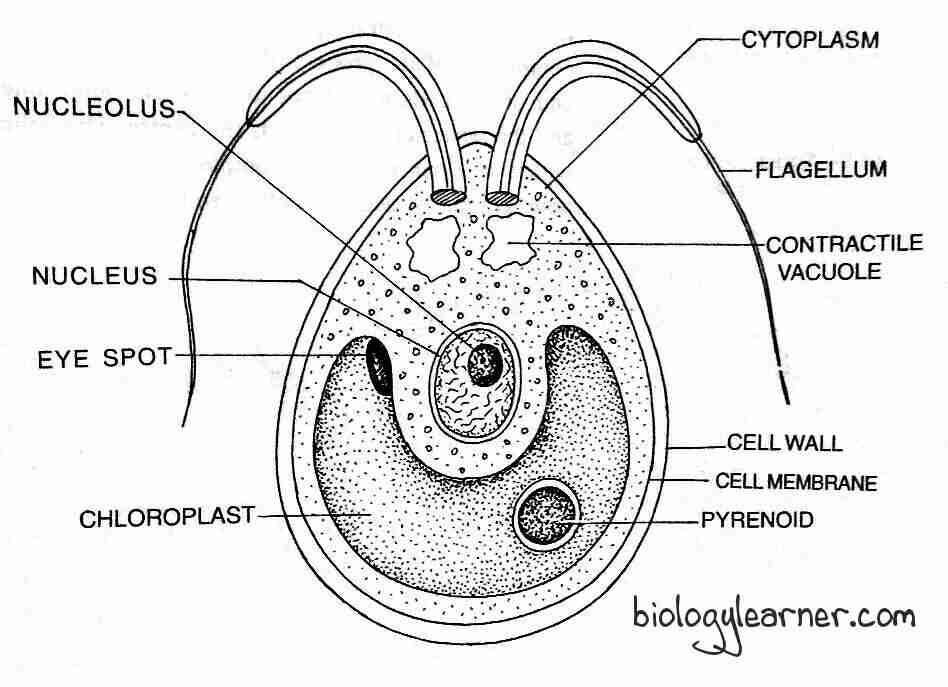
The cell has a thick cell wall differentiated into an outer firm and an inner gelatinous layer. The protoplasm of the cell is embedded within a plasma membrane.
In the cytoplasm, each cell contains a cup-shaped chloroplast with one or more pyrenoids, an eyespot, 2–6 contractile vacuoles, and a single nucleus.
- The contractile vacuoles are found near the surface of the protoplast.
- The cup-shaped or curved plate-like chloroplast contains photosynthetic pigments (chlorophyll a, chlorophyll b, carotenoids, and xanthophylls) characteristic of green algae. There are 1 or 2 pyrenoids associated with the chloroplast.
- The single nucleus is situated in the center portion of the cytoplasm inside the cavity of the cup-shaped chloroplast. It is connected with neuromotor apparatus consisting of blepharoplast, rhizoplast, and centromere.
- A single eyespot is present at the anterior end.
The cells of the anterior region of the coenobium possess larger eyespots than those of the posterior region, indicating the clear polarity of the coenobium.
The cells are interconnected to each other through cytoplasmic strands. In some species of Volvox, such as V. tertius and V. mononae, the cytoplasmic strands are absent. The cytoplasmic strands connect the cells through the mucilage. The central region of the coenobium is generally hollow but, in some species, it is filled with water (V. globator) or gelatinous materials (V. aureus).
Each cell of coenobium independently carries out its own photosynthesis, respiration, and excretion. The movement of the coenobium occurs through coordinating flagellar movement.
Reproduction in Volvox
Volvox reproduces by both asexual and sexual methods. Asexual reproduction occurs under favorable conditions during spring and early summer. Whereas sexual reproduction takes place under unfavorable conditions towards the end of the summer months.
In Volvox, generally, the cells of the coenobium’s posterior end take part in reproduction. These reproductive cells are recognizable by their larger size, prominent nucleus, dense granular cytoplasmic content, more pyrenoids, and lack of flagella.
Asexual Reproduction
During the summer, asexual reproduction occurs very rapidly. Some cells from the posterior side of the coenobium become reproductive. These cells enlarge up to 10 times, withdraw their flagella, and become more or less round. These cells are referred to as gonidia, or parthenogonidia, or autocolony initials.
The number of pyrenoids increases as the gonidia lose their eyespots. The gonidia are enclosed by a gelatinous sac and are pushed inside the colony.
Development of the Daughter Colony
The gonidium undergoes repeated divisions about 15 or more times and can produce more than 3,200 cells. Those cells ultimately form a colony.
Initially, the gonidium undergoes longitudinal division into the plane of the colony and forms 2 cells. The second division is also longitudinal but at a right angle to the first division, forming four cells. These 4 cells again divide longitudinally to form 8 cells, of which 4 cells are central and 4 are peripheral.

These 8 cells are arranged in such a manner that their concave inner surface face toward the outer side of the colony to form a curved plate-like structure. This stage is called the plakea stage, or the cruciate plate stage. Each of these 8 cells divided by longitudinal division forms a 16-celled stage.
At this stage, the cells are arranged in the form of a hollow sphere with an opening towards the exterior side, called a phialopore (a small aperture). At this stage, the cells continue to divide longitudinally until the number of cells reaches the number specified for a particular species.
The cells are naked and in close contact with one another. The anterior end of the cells is directed towards the center and the posterior end towards the outside. This group of cells then undergoes inversion through the phialopore, resulting in the normal pattern of the colony being achieved.
Inversion of Colony
The inversion of the colony begins with the formation of a constriction opposite to Phialopore. The cells of the posterior end, along with constriction, are pushed inside the sphere. At the same time, the phialopore becomes enlarged, through which the lower part comes out and the edges of the phialopore hang backward.
With the help of inversion, the anterior end of the cells changes its position from inner to outer, and the position of the phialopore becomes reversed, i.e., it changes its position from outer to inner. The phialopore gradually closes, forming a completely hollow sphere.
After completion of the inversion, the cells generate a gelatinous cell wall, flagella, and an eyespot. As a result, the daughter colony develops. Many such colonies may form in a coenobium. The daughter colonies initially remain attached to the gelatinized wall of the mother coenobium, swimming freely inside the gelatinous matrix.
Later on, the daughter colonies are released into the water after the rupture or disintegration of the mother coenobium.
In some species, such as V. carteri and V. africanus, daughter colonies of 2-4 generations may remain inside the mother coenobium.
Sexual Reproduction
In Volvox, sexual reproduction is of the oogamous type. Some species are homothallic or monoecious (e.g., V. globator), where the antheridia and oogonia develop in the same colony. Other species (e.g., V. rouseletii) are heterothallic or dioecious, as antheridia and oogonia develop in separate colonies.
The majority of homothallic species are of the protandrous type, i.e., antheridia develop and mature earlier than oogonium. Some species are protogynous types, i.e., oogonia develop and mature before antheridia. V. aureus is usually heterothallic, but it can also be homothallic.

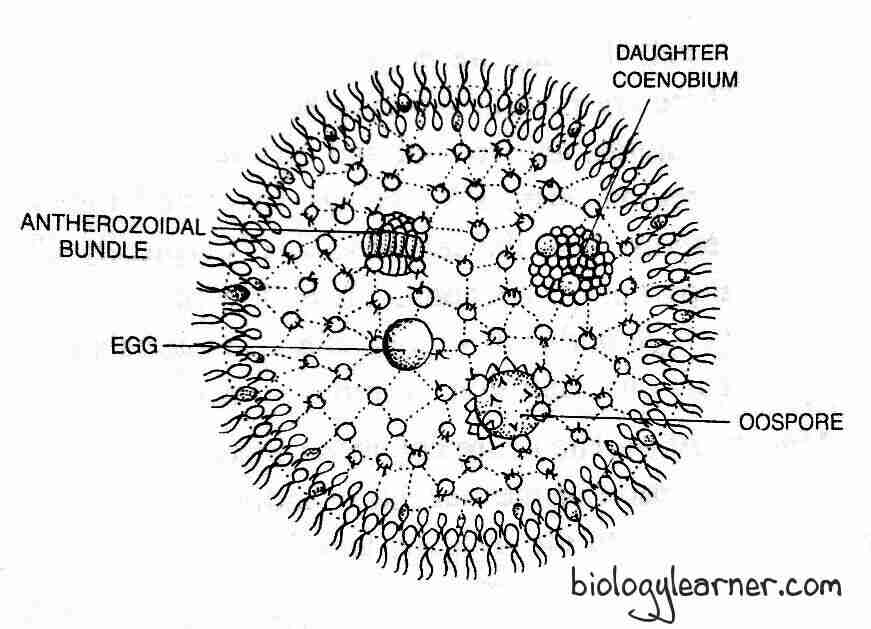
Some specialized cells at the posterior end of the coenobium enlarge in size, withdraw their flagella and develop into reproductive bodies called gametangia. The male gametangia are called antheridia or androgonidia, and the female gametangia are called oogonia or gynogonidia.
Antheridium
The development of the antheridium begins with the formation of the antheridial initial or androgonidial cell, mainly at the posterior end of the coenobium. Similar to gonidia, the initial cells enlarge, lose flagella, the protoplasm becomes dense, and the nucleus becomes larger. The antheridial initial shifts inside the cavity and remains connected to other vegetative cells through cytoplasmic strands.
The protoplast of the antheridial initial undergoes repeated longitudinal divisions, similar to the asexual stage, and forms approximately 64-128 cells (though the number varies from 16-512 depending on the species). Like in the asexual stage, the cells remain in a plate-like structure or are grouped into a hollow sphere and then undergo inversion in which the anterior side of the cells faces the outer side. Each cell develops into an antherozoid or spermatozoid.
The antherozoid is a spindle-shaped, bi-flagellated, elongated, pale yellow or green-colored structure. It contains two contractile vacuoles, a nucleus, a cup-shaped chloroplast with pyrenoids, and an eyespot. Individually or sometimes in groups, the antherozoids are liberated from the antheridium by rupturing the antheridial wall.

Oogonium
The development of the oogonium begins with the formation of the oogonium initial or gynogonidial cell (single vegetative cell) at the posterior end of the coenobium. The oogonial initial enlarges, loses its flagella, the eyespot disappears, and develops into an oogonium. In oogonium, the protoplasm and nucleus become dense.
The oogonium is an enlarged, more or less flask-shaped structure. Without undergoing any division, the entire protoplast of an oogonium forms a uninucleate egg or oosphere or female gametophyte.
The oosphere is spherical, uninucleate, non-flagellated, green in color, and contains a parietal chloroplast. It possesses a large amount of reserve food and many pyrenoids.
The beak of the flask-shaped oogonium opens towards the outer surface of the coenobium and functions as a receptive spot.
Fertilization
After liberation from antheridium, the antherozoids swim freely on the surface of the water. Due to the chemotactic stimulation, they get attracted to the surface of oogonium.
The antherozoids secrete a proteolytic enzyme. With the help of the proteolytic enzyme, some antherozoids enter the egg by breaking the oogonial wall. Only one antherozoid fertilizes the egg.
After fertilization (plasmogamy and karyogamy), a diploid zygote or oospore is formed.
Zygote
The zygote secretes a three-layered (exospore, mesospore, and endospore) thick wall. The outer exospore is quite thick. It may be smooth (V. monanae, V. globator, etc.) or spiny (V. spermatophora). Whereas the exospore and endospore are relatively thin and smooth.
The wall accumulates the pigment haematochrome and it becomes red in color.
The zygote is liberated by the disintegration of the gelatinous matrix of the parent colony and remains dormant for a long period.
Germination of Zygote
Under favorable conditions, at the end of the resting period, the zygospore begins to germinate. During germination, the diploid zygote nucleus (2n) undergoes meiosis to form 4 haploid cells.
The inner layer of the zygote emerges as a vesicle after the outer two layers of the zygote break. The four haploid cells migrate with the vesicle.
The development of the zygote varies among different Volvox species
- In V. minor and V. aureus, the protoplasm of the zygote divides repeatedly and forms a new colony, similar to asexual reproduction.
- In V. campensis, the protoplast of the zygote divides to produce numerous biflagellate zoospores. Only one zoospore survives, while all the others disintegrate. The surviving one comes out of the vesicle and, by repeated mitotic division, it forms a new colony.
- In V. rouseletti, the protoplast of the zygote divides to form 4 zoospores, but only one survives. The outer wall of the zygote bursts, and the inner wall comes out in the form of a vesicle containing a single biflagellate meiospore. The meiospore is liberated by breaking the inner wall. As in asexual reproduction, the protoplast of the meiospore divides and forms a colony.

Botany 1st year