Sphagnum, a genus of mosses (a group of non-vascular, flowerless plants) in the order Sphagnales, is widely distributed all over the world.
- It belongs to the family Sphagnaceae.
- Sphagnum is commonly known as peat moss, bog moss, or turf moss due to its ecological importance in the development of peat or bog.
| Division: | Bryophyta |
| Class: | Bryopsida |
| Sub-Class: | Sphagnidae |
| Order: | Sphagnales |
| Family: | Sphagnaceae |
| Genus: | Sphagnum |
Distribution of Sphagnum
- Sphagnum is a genus of about 336 species. It is found all over the world.
- The species of Sphagnum are perennial plants and grow in dense masses, forming cushion-like structures in swamps, ponds, and wet rocky slopes.
- Sphagnum often completely covers the surface of water, transforming it into bogs of peat lands.
- Some of the common Sphagnum species are: Sphagnum ovatum, S. teres, S. nemoreum, S. robustum, etc.
Gametophyte of Sphagnum
The plant body of Sphagnum is gametophytic. It consists of an erect, radial, leafy stem.

Morphology of Sphagnum
The gametophyte of Sphagnum shows two distinct stages of development: protomena and gametophore. The protonema represents the juvenile stage, while the gametophore is the leafy stage of the mature gametophyte.
Protonema
The protonema is a haploid, irregularly lobed, algal thallus-like structure. It is produced by the germination of the spore. The protonema is attached to the substratum by multicellular rhizoids.
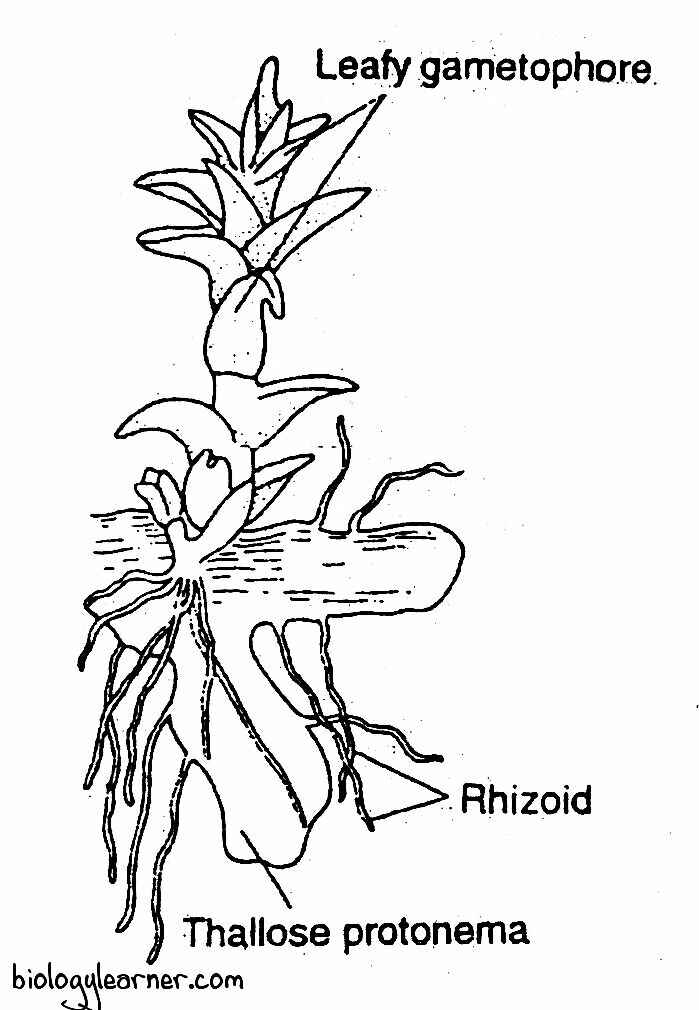
Gametophore
The leafy gametophore arises from the margin of the protonema.
In the young gametophore, a few rhizoids can be seen at the base of the stem. But the mature gametophore does not possess rhizoids (the rhizoids soon disappear as the gametophore grows).
The gametophore (mature gametophyte) of Sphagnum is perennial. It is differentiated into two parts: the axis, or stem, and the leaves.
Axis or Stem
The main axis or stem is about 30 cm or more in height. It is weak and fragile at the young stage but becomes erect and stout at maturity.
The axis is profusely branched, and the branching is lateral. The branches usually arise in fascicles or tufts from the axil of every fourth leaf on the main axis. There are 3 to 8 branches in a tuft.
At the apex of the stem, the branches are short, stout, and of limited growth. These branches are closely clustered together, forming a compact head called the coma or comal tuft.
The coma formation occurs due to the condensed growth of apical internodes present at the apex. As the apical bud grows, these short branches elongate, become normal branches, and are displaced downward.
All the branches in the tufts are similar in structure in submerged aquatic species (e.g., S. cuspidatum, S. obesum). In terrestrial species, the branches are differentiated into two types: divergent branches and flagelliform or drooping branches.
Divergent Branches
The divergent branches are generally short, stout branches that grow out laterally from the axis. They extend outward in a horizontal position and sometimes slightly upward.
However, these branches usually become arched at the middle, with their distal portions carving downward.

Flagelliform or Dropping Branches
The flagelliform branches are long, slender, loosely arranged, descending branches that drop or hang down, often very close to and around the main axis.
These branches are also known as pendent or de-current branches. They function as water conductors in terrestrial Sphagnum.
Leaves
The leaves are small, thin, sessile, oblong, and have a pointed apex. They are arranged spirally on the stem with a phyllotaxy of 2/5.
Each leaf is attached to the stem or branches by a broad base without a distinct midrib. The margin of the leaf is entire.
The leaves are borne on both the main axis and the side branches. On the branches, the leaves are overlapping (as they are positioned very close to each other). But they are placed a little apart on the main axis.
The leaves on the branches are smaller than those on the stem.
Anatomy of Sphagnum
Axis or Stem Anatomy
In a transverse section, the mature stem of Sphagnum shows three distinct regions: the cortex (hyalodermis), the hadrome (prosenchymatous region), and the medulla (axial cylinder).

Cortex or Hyalodermis
The cortex, or hyalodermis, is the outermost region of the stem. It is only one cell thick in the young stem and branches, but several cells (3 to 6 layers) thick in the older stem.
The thickness of the cortex also varies in different Sphagnum species. In S. subsecundum, it is one cell thick throughout its life. In S. obesum and S. recurvum, the cortex is two to three layers thick, while it is composed of two to five layers of cortical cells in S. squarrosum and S. acutifolium.
Initially, the cortex is composed of small, compactly arranged cells. Later, the cells become empty, hyaline, and large in size as they gradually lose their protoplasmic contents and increase in size. In the mature stem, the cortex thus consists of large, dead hyaline cells.
Based on the nature of the hyaline cells, the genus Sphagnum has often been divided into two sub-genera: Inophloea and Lithoploea.
In the sub-genus Inophloea (e.g., S. palustre), the hyaline cells develop large oval pores and spiral thickenings on their walls and become spongy or porous in nature. But they lack pores or spiral thickenings in the sub-genus Lithoploea.

The cortex of the stem can absorb water by capillary action and store the water within the dead hyaline cells.
In a few species, such as S. molluscum and S. tenellum, some cortical cells of the side branches enlarge peculiarly and become retort or flask-shaped, known as retort cells. Each retort cell has a curved neck (turning outward from the stem) with an opening at the distal end.
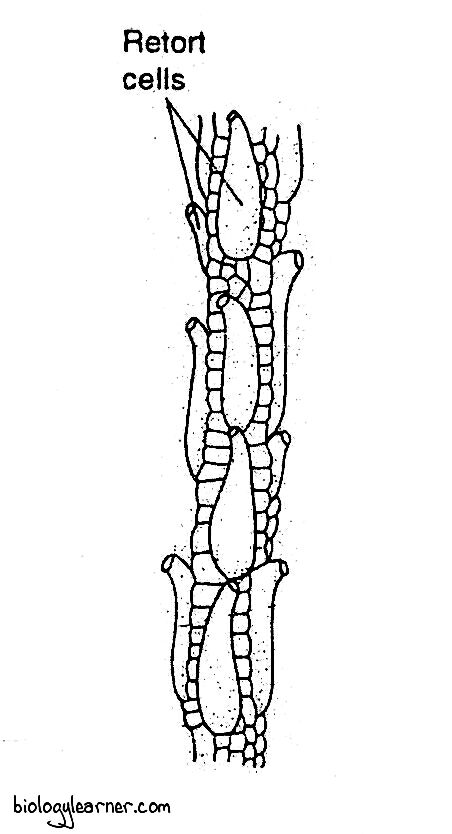
The retort cells accumulate water and are inhabited by small, microscopic animals.
Hadrome or Prosenchymatous Region
The hadrome or prosenchymatous region lies between the cortex and the medulla. It is 4-6 layers thick and consists of small, elongated, thick-walled prosenchymatous cells.
The hadrome gives mechanical support to the axis.
Medulla or Axial Cylinder
At the centre of the stem is the medulla, or axial cylinder. It is composed of small, thin-walled, colourless, vertically elongated collenchymatous cells.

The medulla is somewhat like the pith of the higher plants and serves as the storage region.
Structure of Leaf
Structurally, the leaf of Sphagnum is one cell thick. It consists of much elongated cells. In young leaves, the cells are square or rectangular and are of uniform size.
In a cross-section, the mature leaf appears as a moniliform or bead-like structure. The bead is composed of two types of cells: the ordinary type capillary cells and the green photosynthetic or assimilatory cells. These two types of cells regularly alternate with each other to form a reticulated pattern.
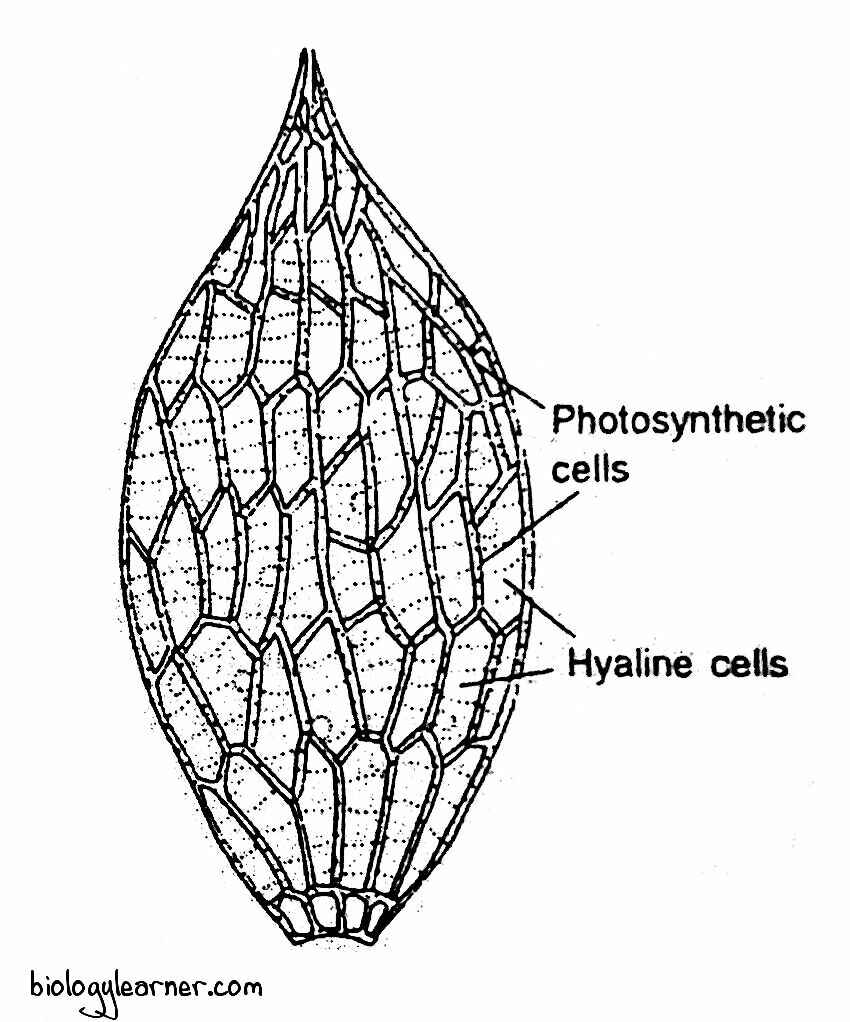
The capillary cells are large, more or less rhomboidal in shape, and become hyaline or colourless by losing their protoplasmic contents. They often form oval or circular pores and spiral thickenings of bands on their walls.
The hyaline cells are capable of absorbing and retaining water. So, rhizoids are not necessary in mature gametophytes.
The assimilatory cells are small, green, living cells. They are biconvex or triangular in shape and contain many discoid chloroplasts.
The assimilatory cells lie between the hyaline cells, forming a network.
According to the species, the assimilatory cells are variable in form and position.
In S. acutifolium, the assimilatory cells are triangular, and the base of the triangle is directed towards the upper surface of the leaf. The large hyaline cells bulge towards the lower surface.
In S. tenellum, the position of both types of cells is reversed. The base of the triangular assimilatory cells faces the lower surface, while the hyaline cells bulge towards the upper surface of the leaf.
The leaves of S. squarrosum have fusiform or spindle-shaped assimilatory cells. Here the assimilatory cells are hemmed in between the hyaline cells, and their ends reach neither the upper nor the lower surface of the leaf.

Due to the presence of chloroplasts, the assimilatory cells are photosynthetic in function and also called photosynthetic cells.
Reproduction in Sphagnum
Sphagnum reproduces by both vegetative and sexual methods, like other bryophytes.
Vegetative Reproduction
Vegetative reproduction is quite common in Sphagnum. It occurs through the following methods:
Innovation
Innovation is the most common method of vegetative reproduction in Sphagnum.
Sometimes, one of the divergent branches in the axillary cluster becomes robust and grows upward. This long, upright branch shows all the characteristics of the main stem and is called the innovation.
Due to the progressive death and decay of the basal, older part of the main stem, the innovation gets separated from the parent plant. Each innovation grows independently by apical growth and develops into a new plant.
In this way, Sphagnum grows in extensive masses in a particular area and is constantly invading new territories.
Multiplication of Protonemal Branches
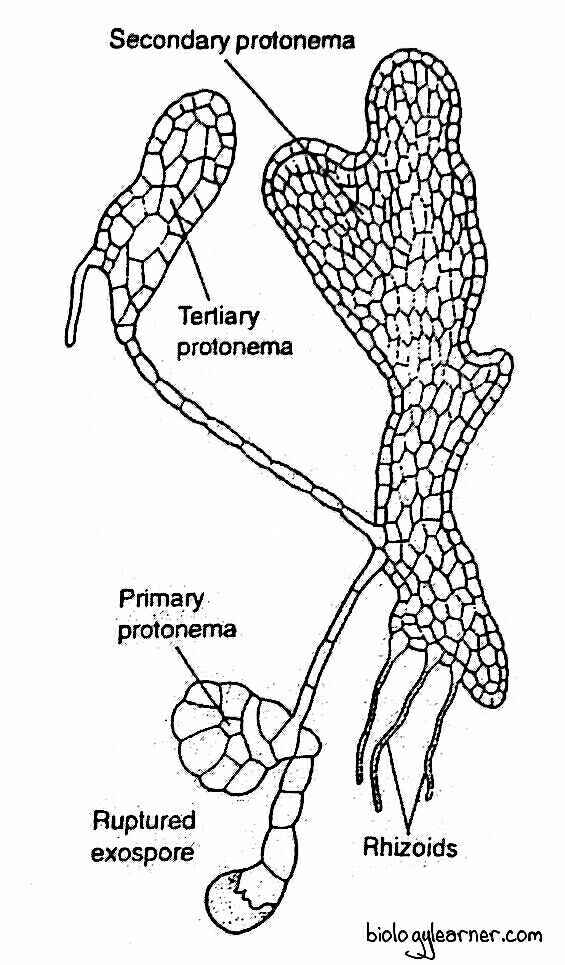
Vegetative reproduction in Sphagnum also occurs through the multiplication of the protonemal branches.
Any marginal cell of the primary protonema may become meristematic. It develops into a green cellular filament.
The apical portion of the filament grows into a flat, thallus-like secondary protonema. From one of the marginal cells in the secondary protonema arises the leafy gametophore.
Regeneration
The growth of new tissues or organs to replace those damaged or lost in injury is called regeneration.
During periods of desiccation, the growth of the Sphagnum is checked because of the suspension of metabolism (respiratory and photosynthetic activities). But the cytoplasm itself shows a high degree of resistance to desiccation.
When water is once again available, the metabolism is resumed, and the normal growth of the plant occurs.
According to Buch (1947), such plants are called pallacuophytes.
Sexual Reproduction
Sexual reproduction in Sphagnum is of the oogamous type. The sex organs are developed only in favourable environmental conditions during the autumn.
The male sex organ is the antheridium, while the archegonium is the female sex organ. The sex organs antheridia or archegonia are produced on special slightly modified branches. These branches are much shorter than the vegetative branches and either occur in the terminal branch cluster (i.e., the comal tuft) or in lower down on the stem.
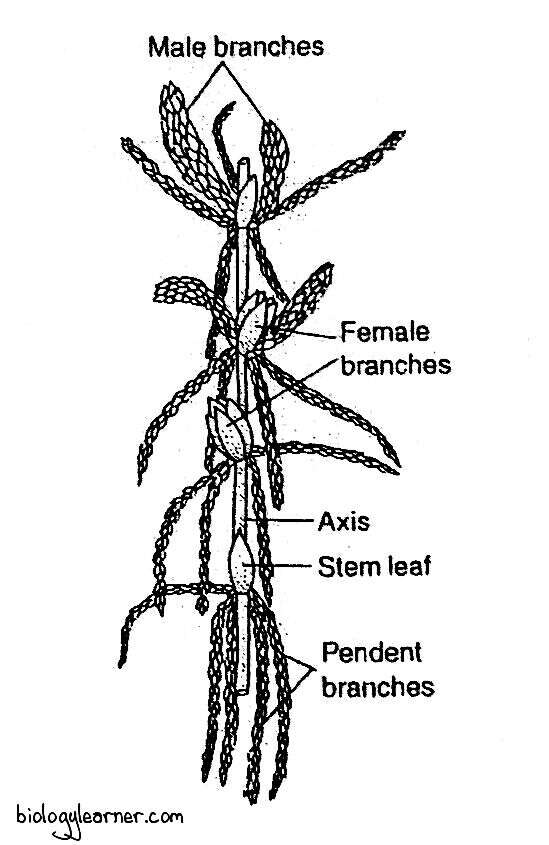
The branches bearing the antheridia are called the antheridial branches, and the branches bearing the archegonia are called the archegonial branches.
Sphagnum may be monoeciuos (sex organs grow on the same plant) or dioecious (sex organs are borne on different plants). In monoecious species, antheridia and archegonium are always borne on separate branches of the same gametophore.
The monoecious species of Sphagnum are protandrous, i.e., the antheridial branches develop first.
Antheridial Branch
The antheridial branches are spindle-shaped, catkin-like lateral branches. These branches are generally shorter but stouter than the vegetative branches.
Antheridial branches develop near the apex of the main shoot. They are densely covered with red, purple, brown, dark green, or yellow leaves, which are usually smaller than the foliage leaves.
Antheridium
The male reproductive organ of Sphagnum is the antheridium. The antheridia occur singly in the axil of each leaf, towards the tip of the antheridial branches.
Development of Antheridium
- In Sphagnum, each antheridium develops from a single superficial cell of the stem called the antheridial initial.
- The antheridial initial grows into a papilla-like outgrowth, and later it becomes a small filament of a few cells.
- The terminal cell of the filament functions as an apical cell. It has two cutting faces.
- The apical cell forms segments in two rows in a regular alternate sequence. In this way, 12 to 15 segments are cut off.
- The last 2–5 segments (i.e., distal cells) form the body of the antheridium. While the lower cells of the row divide vertically and transversely to form the antheridial stalk.
- Each of the distal segments undergoes two successive vertical divisions.
- The first vertical wall divides the segment into two cells of unequal size. The smaller cell is called the first jacket initial.
- The larger cell further divides periclinally into a second jacket initial and a primary androgonial cell.
- The jacket initials divide anticlinally to form the single-layered wall or jacket of the antheridium.
- The primary androgonial cells undergo further divisions and form the androgonial cells.
- The last generation of androgonial cells is called the androcyte mother cells.
- Each androcyte mother cell divides to form two androcytes.
- The androcytes ultimately metamorphose into spirally coiled, biflagellate antherozoids.

Structure of Antheridium
The mature antheridium of Sphagnum consists of a globular body with a long, multicellular stalk. The stalk may be as long as the body of the antheridium and composed of 2-4 vertical rows of cells which are 8–10 cells in length.

The body of the antheridium is covered by a single-layered jacket (antheridial wall) of sterile cells. A very conspicuous layer of cuticle is present on the outer side of the antheridial wall (Manton, 1954).
The sterile jacket encloses a mass of androcyte mother cells. The androcyte mother cell divides to form two androcytes.
Each androcyte metamorphoses into an antherozoid or sperm.
Antherozoids
The antherozoid or sperm is an elongated, unicellular, biflagellate, and spirally coiled structure.
The coiled body remains attached to the posterior end with a cytoplasmic vesicle. The pair of flagella are present at the end, which is cytoplasmic in origin and known as the flagellophore.
Dehiscence of Antheridium
The mature antheridium dehisces with the help of water.
The apical cells of the jacket absorb water and swell up. Due to the turgor pressure, the jacket of antheridium breaks into a number of irregular lobes at the apex, which eventually turn backwards. The antheridium thus opens at its distal end.
A large number of antherozoids come out from the apex of the antheridium and swim freely in the water.
Archegonial Branch
The archegonial branches are very short and green, bud-like. These branches are more or less globular in form.
In many species of Sphagnum, the archegonial branch develops just below the apex of the stem within the comal tuft.
The leaves on the archegonial branches are much larger than the other leaves present on the gametophore. The upper leaves form the perichaetium, which surrounds the archegonia and protects the archegonia from injury.
Archegonium
The female reproductive organ of Sphagnum is archegonium. The archegonia occur either singly or in groups on the apex of the archegonial branches.
Development of Archegonium
- The archegonia of Sphagnum develop on the apex of the female branches (archegonial branches) either singly or in groups.
- The apical cell of an archegonial branch directly functions as an archegonial initial and forms the primary archegonium.
- The other archegonia, which develop from derivatives of the apical cell are called secondary archegonia.
- There are usually three archegonia (the number, however, varies from 2 to 5 in different species) developed on one archegonial branch, of which one primary archegonium is at the apex and two secondary archegonia emerge from the base of the primary archegonium.
- The development of both the primary and secondary archegonia is similar.
- The archegonial initial undergoes a few transverse divisions to form a short filament of 4 to 6 cells.
- The lower cells of the filament form the stalk of the archegonium. While, the terminal cell of the filament enlarges in size. It then divides by three intersecting vertical walls, resulting in the formation of three peripheral jacket initial cells surrounding a single central cell called the primary axial cell.
- The primary axial cell now divides transversely into an upper cover initial and a lower central cell.
- The central cell divides transversely and forms an upper primary neck canal cell and a lower primary venter cell.
- The primary neck canal cell undergoes a series of transverse divisions and forms a row of 8 to 9 neck canal cells.
- The primary ventral cell divides by a single transverse division to form a ventral canal cell and an egg cell.
- The cover initial divides by repeated vertical walls to form a group of eight or more cover cells. These cells, by further divisions, form the upper portion of the archegonial wall or jacket (jacket of neck).
- Subsequently, the jacket initials divide anticlinally and periclinally to form the lower portion of the neck and the venter wall (lower portion of the archegonial wall).
- The basal portion of the neck and the venter undergo further divisions (i.e., periclinally) so that these portions become 2 to 3 layers of cells in thickness.

Structure of Archegonium
The mature archegonium of Sphagnum is a large structure. It has a fairly long stalk.
The archegonium consists of a long twisted neck and a massive venter. The long, twisted neck usually encloses 8 to 9 neck canal cells.

The lower portion of the neck and the venter are 2 to 3 layers thick. The massive venter contains a ventral canal cell and an egg.
Fertilization
Like other bryophytes, the process of fertilization in Sphagnum occurs only in the presence of water.
At maturity, the neck canal cells and the single ventral canal cell in the archegonium disintegrate to form a passage for the antherozoids.
The liberated atherozoids swim freely in the water and reach the archegonium. They enter the archegonial neck through the cover cells due to the chemotactic response and reach up to the egg.
One of the antherozoids fuses with the egg and forms a diploid zygote.
Sporophyte of Sphagnum
In Sphagnum, the sporophytic phase is the direct result of the sexual process and begins with the formation of the diploid zygote after fertilization. The diploid zygote is the first cell of the sporophyte.

Development of Sporophyte
- The sporophyte develops from only one archegonium in the archegonial cluster. The other archegonia persist for some time and degenerate gradually.
- The zygote enlarges and secretes a wall around itself. It divides first by a transverse wall to form an outer epibasal cell and an inner hypobasal cell. Both cells are nearly equal.
- Further divisions also take place transversely in both the epibasal cell and the hypobasal cell until a short filament of 6–12 cells is formed.
- The upper 3 to 4 cells of the filament develop into the capsule, while the lower portion of the filament forms a parenchymatous bulbous foot and a short neck-like, inconspicuous seta.
- Each cell of the upper portion is divided by two successive vertical divisions at right angles to each other to form a quadrant of four cells.
- The cells of the quadrant now divide periclinally to form a central mass of cells called the endothecium, surrounded by an outer layer of amphithecium.
- The cells of the endothecium form the columella, a central sterile part of the capsule.
- The amphithecium divides periclinally into an outer sterile layer of jacket initials and an inner fertile layer of sporogenous cells.
- The cells of the jacket initials divide by periclinal divisions and form a four to six layered capsule wall.
- The outermost layer of the capsule wall is called the epidermis. It has rudimentary, nonfunctional stomata.
- The sporogenous cells later form the dome-shaped archesporium, which overarches the columella.
- The archesporium may extend from two to four layers in thickness.
- The cells of the last cell generation of the sporogenous tissue are all fertile and function as spore mother cells.
- Each of the spore mother cells divides reductionally (i.e., meiotic division) to form four haploid spores.
- The spores are encircled within a spore sac developed from the surrounding sterile tissue.

Structure of Sporophyte
The sporophyte of Sphagnum is differentiated into a bulbous foot and a more or less spherical capsule. The foot and the capsule are connected by a short, narrow, neck-like inconspicuous seta.
The sporophyte is elevated on a short, cylindrical, leafless stalk called the pseudopodium. It develops from the apex of the archegonial branch (gmatophyte) after fertilization, especially when the spores are mature. The pseudopodium increases in length and elevates the capsule far above the perichaetial leaves to facilitate the spreading of spores.

The whole sporophyte is covered by the calyptra. The basal portion of the calyptra and the pseudopodia together constitute a sac-like structure known as the vaginula. The vaginula covers the foot of the sporophyte.
Foot
The foot is a swollen, bulbous structure composed of many parenchymatous cells. It is deeply embedded in the pseudopodium of the female branch.
The foot functions as a haustorium and absorbs nutrients from the gametophyte for the developing sporophyte.
Capsule
The capsule is a small, spherical structure. It is dark brown or black in colour at maturity.
In a longitudinal section, the capsule shows an outer jacket layer and a central columella, which is over arched by a relatively thin dome-shaped spore sac containing spores. The columella is composed of numerous sterile cells.
The capsule wall or jacket is 4-6 layers thick. The outermost layer of the capsule wall is the epidermis. It consists of compactly arranged cuticularized chlorophyllous cells. Many rudimentary, nonfunctional stomata are present between the epidermal cells. Each of the stomata consists of two guard cells without any stomatal aperture.
At the top of the capsule is the circular, convex disc-shaped operculum or lid. The operculum is separated from the rest of the capsule region by a ring-like groove of thin-walled cells known as the annulus.
Dehiscence of the Capsule and Dispersal of Spores
In Sphagnum, the dehiscence of the capsule takes place on a bright sunny day by an explosive mechanism (i.e., a process of air gun mechanism).
The capsule wall and the columella dry up and shrivel under the influence of the sun. The cells of the columella break off and form a large air cavity below the spore sac. Air enters the cavity through the rudimentary stomata.
The spherical capsule gradually becomes cylindrical. Due to the change in shape of the capsule, the imprisoned air in the lower half of the capsule is compressed and held under considerable pressure (4-6 atmospheres).
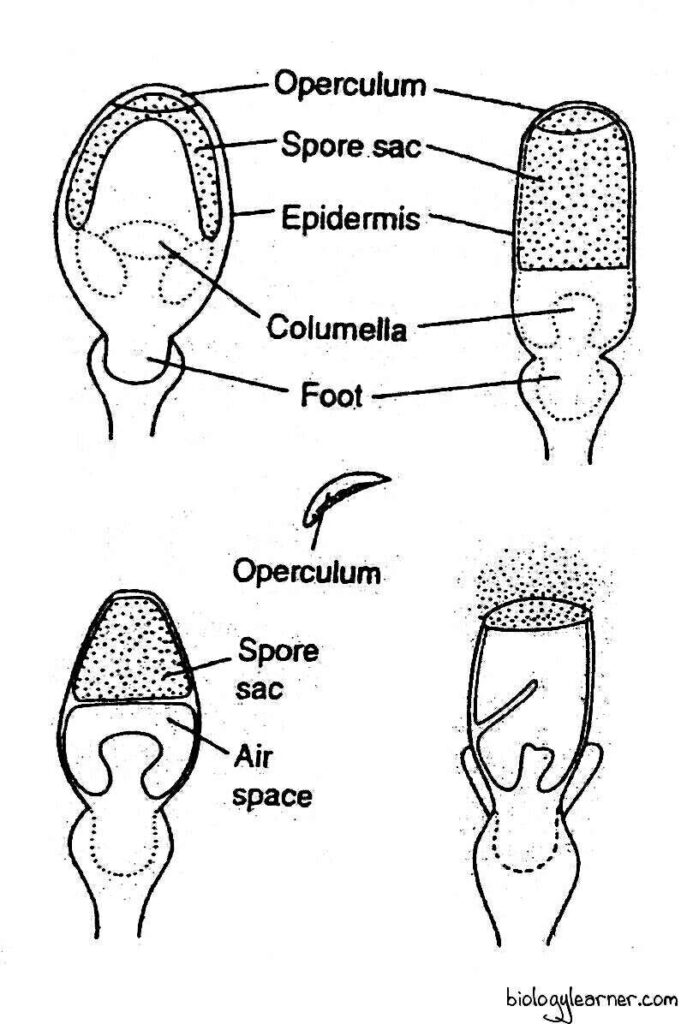
As the capsule wall shrinks, the thick operculum cells resist shrinkage. A difference in tension is thus created, and it puts a strain on the thin-walled annulus cells, which finally rupture under the mounting pressure of imprisoned air.
Eventually, the loosened, small operculum is hurled off with explosive force. The imprisoned air is suddenly released, and with it, the cloud of dry and yellow or orange powdery spores is blown to a height of several centimetres in the atmosphere.
The process of spore dispersal in Sphagnum is, therefore, also called the air gun mechanism (Ingold, 1939).
Spore
The spores are haploid, uninucleate, and yellow to orange in colour. They are tetrahedral in shape with a prominent triradiate mark.
Each spore is surrounded by two wall layers: the outer exine and the inner intine. The exine is ornamented, while the intine is smooth and thin.
A tiny mass of cytoplasm is present inside the spore wall. It contains one or more large oil droplets as reserve food.
Germination of Spores and Formation of the Young Gametophyte in Sphagnum
Spore is the first cell of the gametophytic generation. Each spore germinates within a few days under suitable environmental conditions or may remain viable for six months.
After falling on a suitable substratum, the spore absorbs moisture from the substratum and increases in size. The exine ruptures at the triradiate mark, and the intine comes out as a small germ tube.

The germ tube divides by transverse walls to form a short green filament of 2-4 cells. At this stage, the terminal cell of the filament develops two cutting faces and functions as an apical cell.
The apical cell cuts segments alternately on the right and left. As a result, a green plate-like structure is formed called the primary protonema.
During further development, the apical cell loses activity, and some marginal cells of the protonema become active. These cells undergo anticlinal and periclinal divisions and form irregularly lobed, flat, thallus-like secondary protonema with rhizoids and leaf buds.
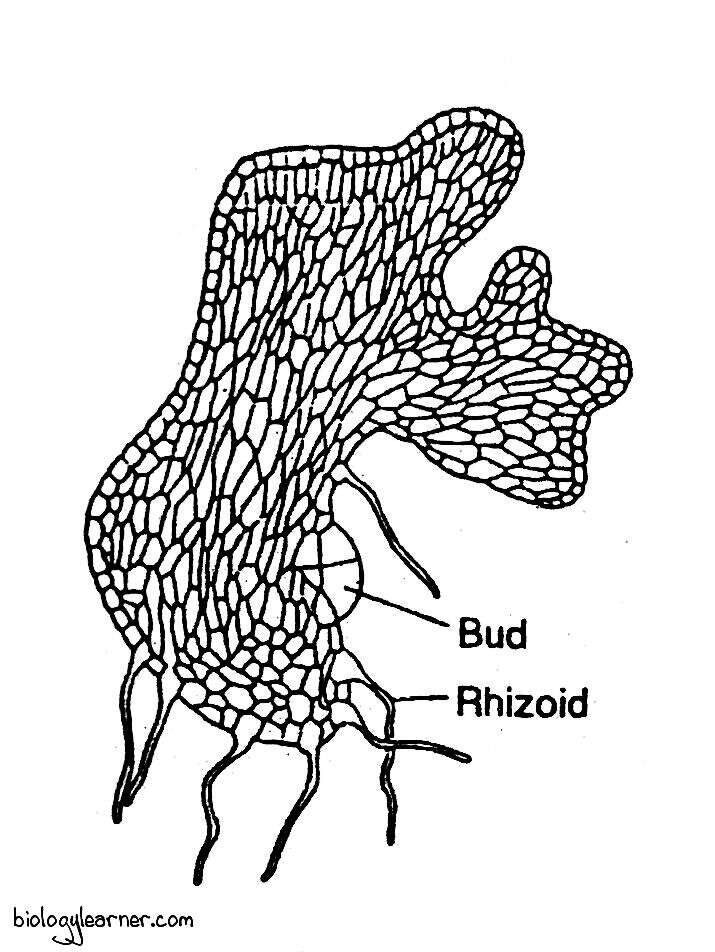
The rhizoids are multicellular, colourless, unbranched, and possess oblique septa between the cells. They anchor the protonema in the substratum.
The leaf bud is produced from one of the marginal cells near the base of the primary protonema. Each bud grows into an erect, leafy gametophore.
Normally, one leafy gametophore is developed from one single protonema in Sphagnum.

Thanks for your article.
Excellent!
Nice post.
Good Information. Thanks