Bryophytes are small, non-vascular land plants, which have multicellular jacketed sex organs.
The defining features of bryophytes are that their life cycle features alternating haploid and diploid generations with a dominant, branched gametophyte stage.
Introduction
Bryophytes are quite soft, and therefore, they lack fossil records. Also, no intermediate forms are available. Hence, the question of the origin of bryophytes has been debated at length.
All the views are based on the evidence under the following three heads:
- Evidence from comparative morphology of the living plants
- Evidence from ontogeny of the living plants
- Evidence-based on analogies with the living plants of other groups
Bryologists are divided into two schools of thought on the origin of bryophytes. One school of thought supports the Algal hypothesis of the origin of bryophytes and the other supports the Pteridophytean hypothesis of the origin of bryophytes.
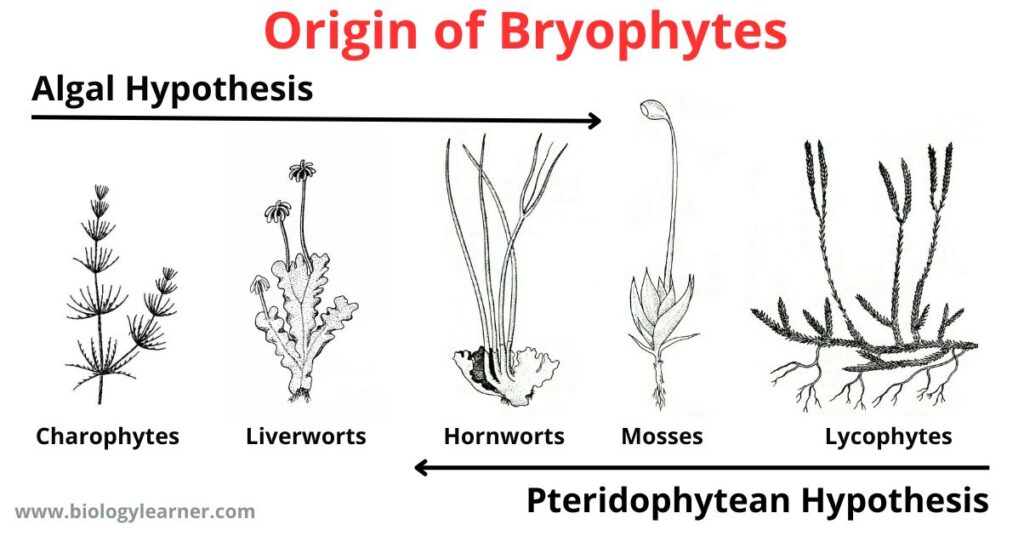
Algal Hypothesis
This view has been supported by Church (1919), Campbell (1940), Frye & Clark (1937-45), Fritsch (1945), and others.
According to the Algal hypothesis, the Archegoniatae ( bryophytes and pteridophytes) were derived from algae. The archegoniatae in turn gave rise to Spermatophyta (seed plants).
It is assumed that bryophytes sprang from such semi-terrestrial algal ancestors in which, due to the scarcity of an external aquatic environment, the fertilization was dependent on waters (rains or dew). As a result, the sporophyte assumed viable form zygote and adapted to the land habits gradually.
The occurrence of cytological alternation of generations in algae and bryophytes also supports this view.
Affinities with Algae
- Amphibian nature.
- Lack of vascular tissues in the plant body (thallus).
- Autotrophic mode of nutrition.
- Prominent gametophytic life cycle.
- Presence of flagellated spermatozoid.
- Water is essential for fertilization.
Church’s View (1919)
Church postulated his theory of the origin of bryophytes from algae on the following assumption:
- The primitive Hepatics, like algae, had morphologically similar sporophytes and gametophytes.
- A terrestrial plant had descended probably from submerged aquatic species of green algae in which cytological alternations were already attained and also remained effective through migration to land plants.
- Somatic equipment of Phaeophyta such as in the multiseptate organization, differentiation into massive main axis and lateral branches, dichotomy, conducting tissues, etc.
Campbell’s View (1940)
According to Campbell, bryophytes have descended directly from Chara. His view is based on the close similarity of the sex organs of Chara and Bryophytes. The sex organs of Chara have a sterile enveloping jacket like the archegonium of bryophytes.
Frye & Clark’s View (1937-45)
Frye and Clark held that both Chara and ancestral primitive hepatics arose on parallel lines from ancient algae, from such primitive hepatics present-day bryophytes (hepatics) sprung.
The primitive ancient algae were supposed to have filamentous gametophytes with apical growth and hairy sperm cells constituting the male filament and enlarged egg cells in a branch that were surrounded by sterile branches. The primitive ancestral hepatics from which present-day hepatics were derived, had dorsiventral body coherent branches, dorsal sex organs, formation of an archegonial jacket, coherence of filaments, etc.
In these primitive hepatics, the sporogenous mass of diploid cells was formed after fertilization, and all the cells except the cells of the jacket layer underwent reduction division to form haploid spores.
Smith’s View (1955)
Smith supposed that Bryophyta has originated from Ulotrichales of Chlorophyta (green algae). The primitive bryophytes, similar to many present-day dry species, probably came into existence when there was plenty of water in the soil.
Thus there was little development of water-absorbing tissues (except for certain cells on the dorsal surface) growing as rhizoids. The abundance of moisture corresponds to the method of gametic fusion as in algae.
Pteridophytean Hypothesis
According to the Pteridophytean hypothesis, the bryophytes have been descended from pteridophytes by means of reduction.
The scientists formulated their argument on the basis of the following two features:
- Close similarity between the sex organs of the two groups.
- The resemblance between sporogonium of Anthocerose, Sphagnum, and terminal sporangium of fossil pteridophytes: Sporogonites and Horneophyton.
This second view, the Pteridophytean hypothesis of the origin of bryophytes has been strongly suggested by many workers such as Kashyap (1919), Christensen (1954), Andrews (1960), Proskauer (1960), etc.
In fact, this theory is a regressive theory that states that the evolution of simple forms has taken place from complex forms due to a reduction in histological complexity. Rather it is not always that complex forms have been derived from simplest forms by elaboration.
Affinities with Pteridophytes
- Distinct and well-defined heteromorphic alternation of generations in the life history.
- Methods of reproduction.
- The origin and the formation of spores from spore mother cells.
- The similarity of their pigments.
- Structure of cell wall.
Kashyap’s View (1919)
According to Kashyap, the hepatics may have arisen from the pteridophytes as there are several features common to both pteridophytes and bryophytes like;
- Thallus structure (thalloid gametophyte).
- The similar structure of sex organs.
- Methods of spore formation.
Further, there is evidence that lower forms of hepatics were derived from higher forms. The gametophyte of Lycopodium cernuum (a pteridophyte) and that of Anthoceros erectus (a bryophyte) are identical. So it has been presumed that the evolution of the dorsiventral gametophyte of Marchantia (hepatics) took place probably from the erect radial gametophyte of pteridophyte.
It has been observed in support of the above view that the gametophyte of Equisetum debile resembles, on one hand, that of Marchantia and on the other Lycopodium cernuum.
So, Kashyap postulated the line of evolution as follows:
Lycopodium cernuum→Equisetum debile→Marchantia
Christensen’s View (1954)
Christensen also suggested the pteridophytean origin of bryophytes. According to him, bryophytes have been derived from any one of the following conditions:
- From pteridophytes bearing leaves in the stems of both the gametophyte and sporophyte.
- From leafless pteridophytes like the members of Rhyniaceae.
- Polyphyletically from different types of pteridophytes.
Proskauer’s View (1960)
Proskauer suggested that anthocerotean types of bryophytes have been derived from the Horneophton type of pteridophyte.
