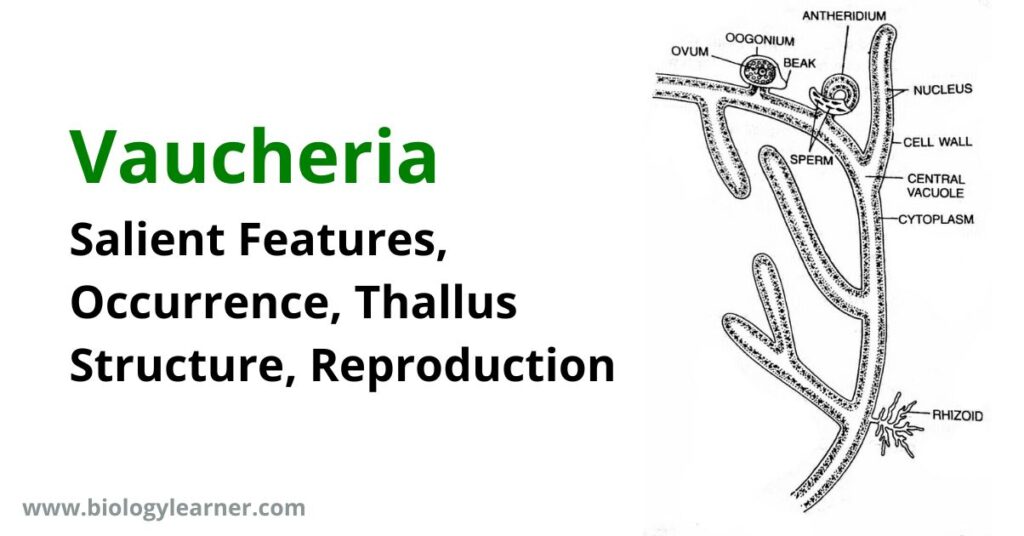
Vaucheria is a freshwater green alga of the class Xanthophyceae.
- Vaucheria is found nearly worldwide, especially in lakes, canals, and wetlands.
- The terrestrial species remain attached to the substratum by colorless rhizoids.
| Class: | Xanthophyceae |
| Order: | Vaucheriales |
| Family: | Vaucheriaceae |
| Genus: | Vaucheria |
Salient Features of Vaucheria
The salient features of Vaucheria are as follows:
- The plant body is filamentous, coenocytic, siphonaceous, and branched.
- Filaments are without septation (not divided into cell walls).
- The plant body is attached to the substratum by colorless branched rhizoids.
- Reserve food is droplets of oil.
- Asexual reproduction takes place by multiflagellate zoospores (coenozoospores). The zoospores can be seen with the naked eye.
- Sexual reproduction is a highly complicated oogamous type.
- The oogonium contains a single large, uninucleate egg, while the antheridium contains many biflagellate antherozoids (sperms).
Occurrence of Vaucheria
Vuacheria is a genus of 54 species, nine of which are found in India. The majority of species are terrestrial and aquatic (freshwater).
Terrestrial species grow in winter in the form of a green and thick layer on damp, moist soil. Aquatic species occur widely in stagnant brackish water (V. litorea) and some marine (V. piloboloides).
Thallus Structure of Vaucheria
The thallus, i.e., the plant body of Vaucheria, is unicellular, filamentous, much-branched, siphonaceous, and coenocytic. The filaments are profusely branched and yellowish-green in color.
The thallus is multinucleate and without any septa (transverse or cross-walls), showing a coenocytic condition. Septa formation takes place during injury or during the development of sex organs. Septate filament is found in Vaucheria pseudohamata.
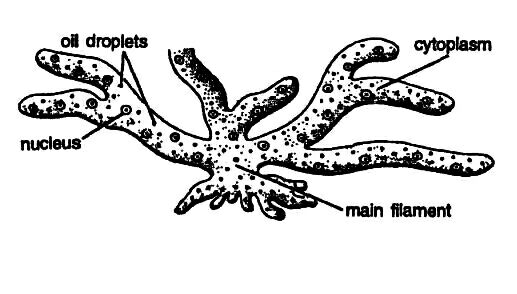
In terrestrial species, the plant body remains attached to the substratum (soil surface) by means of a hapteron-like, colorless, branched structure called the rhizoid. The rhizoids are either absent or poorly developed in free-floating species.
The cell wall of the filamentous body is thin and two-layered. The outer layer is composed of pectose, while the inner layer is made up of cellulose. In the center lies a big vacuole, continuous throughout the length of the filament except at the apical region, which is filled up with cell sap.
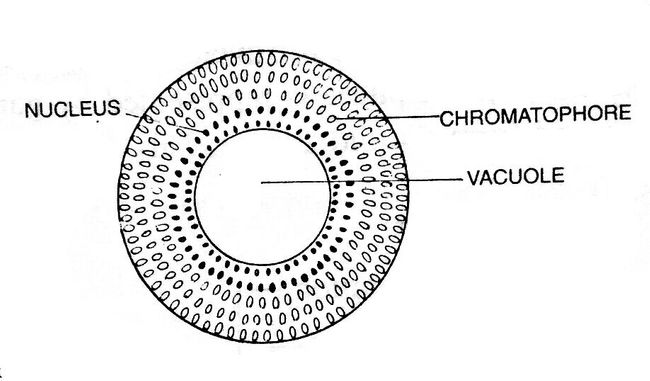
The cytoplasm is present between the vacuole and the cell wall. The cytoplasm contains numerous circular or disc-like chloroplasts, nuclei, and chromatophores. Pyrenoids are usually absent in chloroplasts. Many nuclei are scattered in the cytoplasm near the vacuole.
Small circular or elliptical-shaped chromatophores are also scattered in the cytoplasm. In the cytoplasm, the chromatophores remain toward the periphery.
Vaucheria contains pigments like that of Xanthophyceae such as chlorophyll-a, chlorophyll-e, β-carotene, xanthophylls, etc. Chlorophyll-b is absent. Oil is the reserve food material, and it occurs in the cytoplasm as colorless droplets.
Reproduction in Vaucheria
Vaucheria reproduces in all three methods: vegetative, asexual, and sexual.
Vegetative Reproduction
The vegetative reproduction in Vaucheria takes place by fragmentation. The thallus can break into small fragments due to accidental breakage (mechanical injury or insect bites). Septa are developed in the injured part of the filament.
The broken fragments, which are detached from the thallus, develop into new Vaucheria plants.
Asexual Reproduction
Asexual reproduction in Vaucheria takes place by the formation of various types of asexual spores, such as zoospores, aplanospores, and akinetes.
By Zoospores
The zoospores formation is the most common method of reproduction in aquatic algae. Zoospores are formed singly inside the club-shaped zoosporangium.
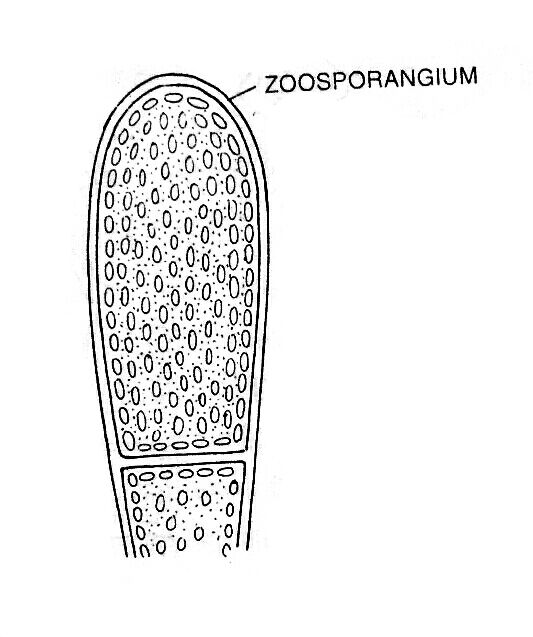
During zoosporangium formation, a club-shaped swelling at the tip of a side branch. A large number of nuclei and chloroplasts along with cytoplasm accumulate towards the apex, as a result, the vacuole becomes reduced in size. At the base of the cytoplasm, a colorless protoplasmic region becomes visible, which is separated from the rest of the cytoplasm of the thallus by a cross-wall and forms apical sporangium.
Consequently, the sporangium becomes very dense, club-shaped, and dark green in color. After some time, the change takes place in the relative positions of chloroplasts and nuclei; the nuclei become peripheral and chloroplasts enter the inner layer of the cytoplasm.
The entire protoplasm of the zoosporangium contracts to form an oval, multinucleate zoospore. During the liberation of zoospores, a terminal aperture develops in the zoosporangium by gelatinization of the wall. The zoospore is liberated through the aperture in the morning hours.
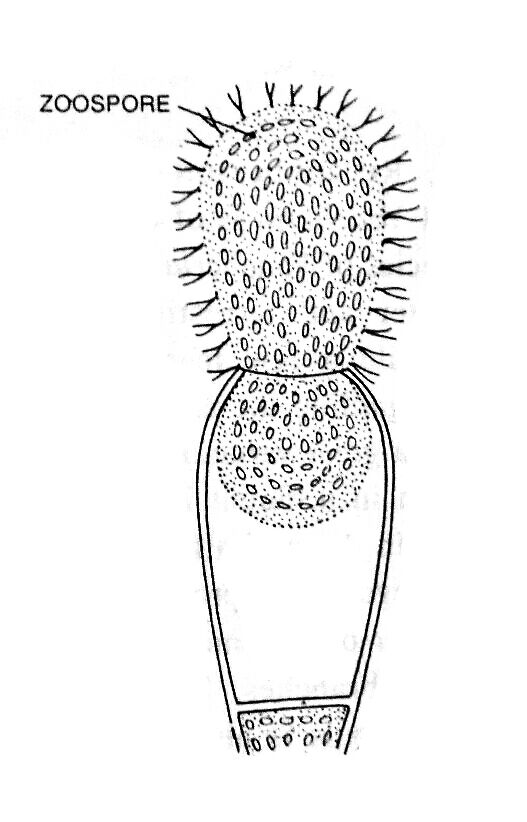
The zoospore is an ovoid, multinuleate, multiflagellate, yellow-green structure. It has a small central vacuole, and many chromatophores are situated between the vacuole and peripheral nuclei. Two flagella (heterokontic and whiplash type) arise opposite each nucleus.
The zoospore of Vaucheria is considered a compound zoospore and is also known as a synzoospore or coenozoospore as a number of zoospores have failed to separate from one another (Greenwood, Manton, and Clarke; 1957).
After liberation, the zoospores swim freely in the water for about 15–30 minutes. They come to rest on the suitable substratum, withdraw their flagella, and secret thin walls.
During germination, the chromatophores of each zoospore move outward and the nuclei inward. The two or more tube-like outgrowths (i.e., germ tubes) develop in opposite directions. One of the germ tubes develops into a new thallus, and the other one elongates to form a colorless lobed hapteron.
By Aplanospores
The aplanospores are generally formed by terrestrial species (V. geminata) and freshwater species. A few marine species (V. piloboloides) also form aplanospores during unfavorable conditions.
The aplanospores are non-motile asexual spores formed singly in special structures called aplanosporangia. The aplanosporangium is club-shaped and is formed at the apical end of the young branch.
Aplanospores are multinucleate and thin-walled. They are liberated by an irregular rupturing of the apical wall. After liberation, the aplanospore germinates into a new thallus.
By Akinetes
Akinetes are formed during unfavorable conditions like droughts, and low temperatures. The akinetes are formed at the terminal part of lateral branches where protoplasm becomes separated into small segments by a thick gelatinous wall. These thick-walled, small, multinucleate segments are called akinetes, hypnospores, or cysts.
The akinetes may divide further into a number of thin-walled bodies called cysts. The chain of these cysts resembles the filament of another alga Gongrosira. Hence, this stage of Vaucheria is called the Gongrosira stage.

Under favorable conditions, the akinetes and cysts germinate into new thalli.
Sexual Reproduction
In Vaucheria, sexual reproduction is of the oogamous type. The male sex organ is antheridium, whereas oogonium is the female sex organ.
Most of the freshwater species are homothallic or monoecious. A few marine species, such as V. dichotoma, V. litorea, and V. mayyanadensis, are heterothallic or dioecious.

Position of Sex Organs
There is a great variation in the arrangement of sex organs in different Vaucheria species and even in different individuals of the same species.
Antheridia and oogonia are borne very close together or next to each other on a single filament or adjacent filaments in homothallic species (e.g., V. sessilis, V. aversa). In heterothallic species (e.g., V. dichotoma), they are borne on separate plants.
In Vaucheria, the position of the sex organs generally has two types: sessile and geminata.
- Sessile type: In this type, the sex organs are formed on the main filament. The male and female sex organs are produced on branches close to each other and are sessile, e.g., V. sessilis.
- Geminata type: In this type, the sex organs are formed on special branches. These branches are short and bear terminal antheridium (a male sex organ) and lateral groups of oogonia (female sex organs), e.g., V. geminata and V. terrestris.
Antheridium
The male sex organ of Vaucheria is the antheridium.
Antheridium Structure and Development
The mature antheridium is a cylindrical, tubular, straight, or strongly curved structure opened by a terminal pore.
In homothallic species, the antheridium is formed at the end of a short lateral branch along with or before the oogonium development. The apical region of the branch becomes densely filled with cytoplasm containing many nuclei and a few chloroplasts. After that the apical region bends like a horn and gets cut off from the lower part, forming a septum (cross wall).
The antheridium grows and becomes a highly curved structure. Its upper portion is the main antheridium, and the lower portion is the stalk. The nuclei of the antheridium aggregate in the center and divide mitotically. Each nucleus, along with some cytoplasm, metamorphosed into a biflagellate antherozoid, or sperm.

The antherozoid is a colorless, spindle-shaped, or pear-shaped structure with two unequal flagella inserted laterally. One flagellum is short and is of the tinsel type, while the other is long and is of the whiplash type.
The mature antherozoids are liberated through a terminal pore developed at the apical region of the antheridium.
Oogonium
The female sex organ of Vaucheria is the oogonium.
Oogonium Structure and Development
The development of oogonium begins simultaneously with an accumulation of a colorless, multinucleate mass of cytoplasm in the main filament near the base of an antheridial branch. This accumulated mass of cytoplasm is called “wanderplasm” (Oltmans, 1895).
The young oogonium, also known as the oogonium initial, is formed by a lateral bulging or outgrowth of the main filament near the wanderplasm. The wanderplasm enters into the oogonial initial. A large amount of cytoplasm and nuclei migrate into the oogonium initial, which gradually enlarges to make it a large globular structure called oogonium.
The oogonium is a sessile or short-stalk structure with a tumid, globose, obovoid, hemispherical, or pyriform shape. The oogonium with a beak at the base gets separated from the main filament by the development of a septum.
The mature oogonium contains a large nucleus at the center. The nucleus of an oogonium is filled with protoplasm to form a single egg, ovum, or oosphere.
Fertilization
During fertilization, both antheridium and oogonium open almost at the same time. The tip of the beak of oogonium gelatinizes and produces an aperture. Simultaneously an aperture is also developed at the apex of the antheridium (the time of antheridium opening may vary from a few minutes to one or two hours).

A small drop of colorless cytoplasm oozes out through the rupturing beak of the oogonium in the form of a twisted mass. In the form of a twisted mass, a small drop of colorless cytoplasm oozes out through the rupturing beak of the oogonium. Many antherozoids liberated from the terminal opening of antheridium get entangled in the drop.
Only one antherozoid enters the oogonium. The nucleus of the antherozoid is smaller than the egg nucleus. The male nucleus i.e., the antherozoid nucleus increases in size and fuses with the egg nucleus to form a diploid zygote.
Zygote or Oospore
After fertilization, the zygote secretes a thick 3–7 layered wall around it, which is now called an oospore. Oil droplets accumulate in the protoplast.
Initially, the oospore remains green, but as the chlorophyll in the oospore breaks down, it becomes red.
For a few months, the oospore remains dormant and stays inside the oogonium.
Germination of Zygote or Oospore
During favorable conditions, the oogonial wall disintegrates, releasing the oospore. The oospore germinates into a new thallus.
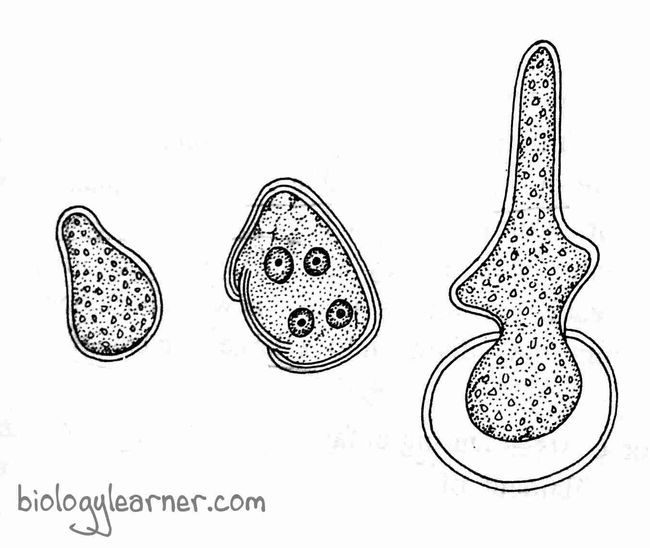
The diploid zygote i.e., oospore undergoes the first meiotic division, followed by several mitotic divisions, and forms the haploid nuclei.
A haploid coenocytic filament comes out of the oospore wall from each nucleus.
Life Cycle of Vaucheria
According to Williams, Vaucheria has a haplontic life cycle, with the oospore being the only diploid structure. The haploid Vaucheria thallus is a branched, cylindrical, or tubular structure showing coenocytic conditions.
The vegetative reproduction in Vaucheria takes place by fragmentation. Asexual reproduction occurs through asexual spores. Aquatic species produce zoospores while terrestrial species produce aplanospores.
Sexual reproduction is an advanced oogamous type. The male and female sex organs are antheridia and oogonia. The majority of the species are homothallic, but some are heterothallic.
A diploid zygote, or oospore, is formed after fertilisation. The oospore goes through a period of rest before germination. During germination, the oospore undergoes reduction division, resulting in the formation of a haploid thallus.
