Gnetales is an order of the gymnospermic seed plants. This order is composed of three genera- Gnetum, Ephedra, and Welwitschia. Chamberlain (1935) has placed all three genera in a single-family Gnetaceae. But Markgraf (1951) has been split up the order Gnetales (due to the heterogeneous characters of those three genera) into three families (e.g., Ephedraceae, Welwitschiaceae, and Gnetaceae). Each family with single genus such as Ephedra, Welwitschia, and Gnetum respectively.
All three genera consist of different characters and Gnetum shows connecting link between angiosperms and gymnosperms. The members of the order Gnetales are characterized by the presence of compound staminate strobili, opposite or whorled leaves. The vessels are present in the secondary wood and absence of resin canals.

Distribution of Gnetales
- The members of Gnetales are widely distributed in Central and Western Asia, South America, and Western Africa.
- In India, they are found in the hill regions.
- Examples: The members of Gnetales are Gnetum ula, G. montanum, G. gnemon, G. latifolium, Ephedra foliata, E. intermedia, etc.
Morphology of Gnetales
External Morphology
Stem
- In Ephedra, the stem is branched and green in color. It is fistular and jointed (with nodes).
- Internodes elongate by the activity of the meristematic tissue lying at the base.
- While in Welwitschia, the stem is underground and its height is about 1-2 meters.
- The stem is woody, branched in Gnetum.
- The branches are of two types: dwarf shoots (short branches of limited growth) and long shoots (long branches of unlimited growth).
- Nodes and internodes are present on the dwarf shoots.
Leaves
- The leaves are very small, scally, and inconspicuous in Ephedra.
- They are cyclic, usually, opposite decussate.
- Each scale leaf is provided with a thick midrib with a thin border.
- In Gnetum, leaves are simple, exstipulate, and shortly petioled.
- The leaf blade (lamina) is oval-shaped, large, and entire with pinnately reticulate venation.
- Leaves (about 9 and 10) are arranged in opposite decussate pairs on each branch.
- In Welwitschia, leaves are opposite, decussate, and petiolated. An apical groove is present.
Root
- In Gnetales, roots are normal tap root types.
- They are branched.
Internal Morphology
Stem Anatomy
- A transverse section of the stem shows an epidermis composed of thick-walled cells in Ephedra.
- Cortex is composed of thin-walled, radially elongated chlorenchymatous cells containing abundant chloroplasts and intracellular spaces.
- In Welwitschia, the cortex is made up of spicular cells. Calcium oxalate crystals are present.
- Vascular bundles are enclosed by fibers.
- Reniform bordered pits are present in the xylem.
- While in Gnetum, three types (external, middle, and internal) cortex is present.
- The outer cortex is made up of chlorenchymatous tissues.
- The middle cortex consists of parenchymatous tissues and the internal cortex contains sclerenchymatous tissues.
- The primary vascular bundles vary in number from 20 to 24, they are arranged in the form of a ring.
- Vascular bundles are collateral, open and endarch. Vessels are large and pith is present.
Internal Structure of Root
- In one layer of epiblema is made up of root hairs.
- Cortex is a simple parenchymatous.
- Starch is present in Gnetum roots.
- Spicular cells are found in the cortex of Welwitschia.
- Endodermis and pericycle are present. Xylem is diarch.
- Secondary growth is normal.
Leaf Anatomy
- In Ephedra, leaves do not exhibit well-marked tissue differentiation as they are scaly and very much reduced.
- A layer of palisade lying below the epidermis constitutes the mesophyll.
- Two vascular bundles are embedded in the mesophyll.
- In Welwitschia vascular bundle is covered by a bundle sheath.
- Latex is present.
- Stomata are present on the lower epidermis which is a syndectocheilic type.
- Spongy tissues are made up of chlorenchyma, sclerenchyma, fibrous tissues, and latex tubes.
- In Gnetum, the upper epidermis of the leaf is thickly cuticularised.
- On the lower epidermis, a large number of stomata are present.
- Vascular bundles are collateral and endarch.
- Fibers and latex tubes are present.
Reproduction of Gnetales
All members of Gnetales are mostly dioecious. Flowers are unisexual. Male flowers are represented by microsporophylls and female flowers are represented by megasporophylls.
In Gnetum, male and female flowers are borne in clusters on male and female plants respectively forming cones or strobili.
Male Strobilus
- In Ephedra, male strobili are borne in whorls in the axils of scale leaves at each node of the fertile branch.
- Each strobilus consists of a short axis upon which are borne 2-8 pairs of opposite decussate bracts.
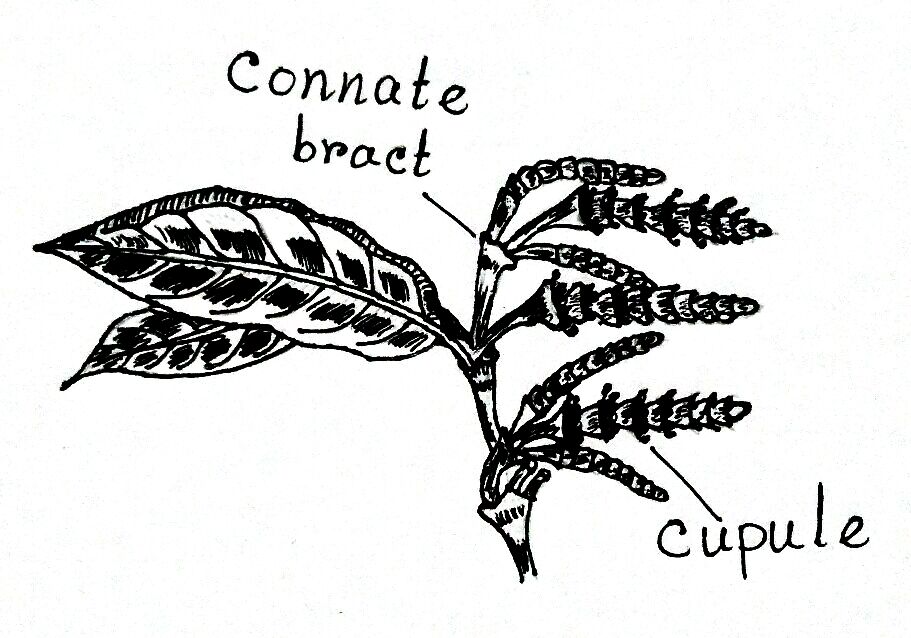
- In Gnetum, the male cones are compact and slender axis-like structures, which are up to 6 cm in length.
- Each cone is generally a penicle that is either solitary and axillary or fascicled at the apex.
- The axis of the strobilus bears opposite and connate bracts.
- Bracts are arranged one above the other to form collars or cupules.
- There are 6-25 cupules are present, each cupule, staminate flowers are arranged in several definite rings, usually 3 to 6 in number.
- Each mature microsporophyll consists of a stalk bearing two unilocular microsporangia (anthers).
- The stalk is invested at the base by a perianth.
- In Gnetum ula, 2-4 anthers are there which are formed from microsporangial initial cells.
- It divides periclinally and anticlinally to form a primary cell wall, tapetal cell, and sporogenous cells.
- Sporogenous cells later differentiate into spore mother cells.
- Microspore mother cell forms microspores (pollen grains) through meiosis.
- The wingless pollen grains are liberated by the longitudinal dehiscence of the anthers.
Female Strobilus
- In Welwitschia, the central axis is surrounded by bracts.
- Each female flower bears one bract towards the upper side and one pair of lateral bracts.
- The ovule is bitegmic. Both integuments form micropyle. Inner integument joins with the nucellus.
- In Gnetum, female flowers bear 4-10 ovules.
- Some ovules enlarge and they have three layers of integuments.
- Only inner integuments form micropyle. While middle and outer integument forms the outer cover.
- The whole structure is covered by perianth.
- In Ephedra, each fertile bract contains one ovule.
- The ovule consists of a distinct nucellus surrounded apparently by inner and outer integuments.
- The tip of the inner integument elongates immediately by the commencement of pollination and forms o short micropylar tube.
- A single hypodermal archesporial cell is differentiated in the nucellus of the ovule.
- The nucellus is bitegmic and is surrounded by 2 envelopes.
- Archesporial cell forms parietal cell and megaspore mother cell which forms 4 haploid megaspores by meiosis.
- The lowermost megaspore cell is the functional megaspore, while the other three facing the micropyle degenerate.
Gametophytes of Gnetales
Male Gametophyte
- Microspore (pollen grain) is the first cell of the male gametophyte.
- The outer exine is thick-walled and the inner intine is thin-walled.
- In Ephedra, the microspore nucleus divides and cuts off the first prothallial cell, which again divides and forms an antheridial initial.
- The antheridial nucleus divides to form a tube nucleus and a body nucleus. The intine elongates forming a pollen tube in the pollen chamber.
- The body nucleus divides to form two male gametes.
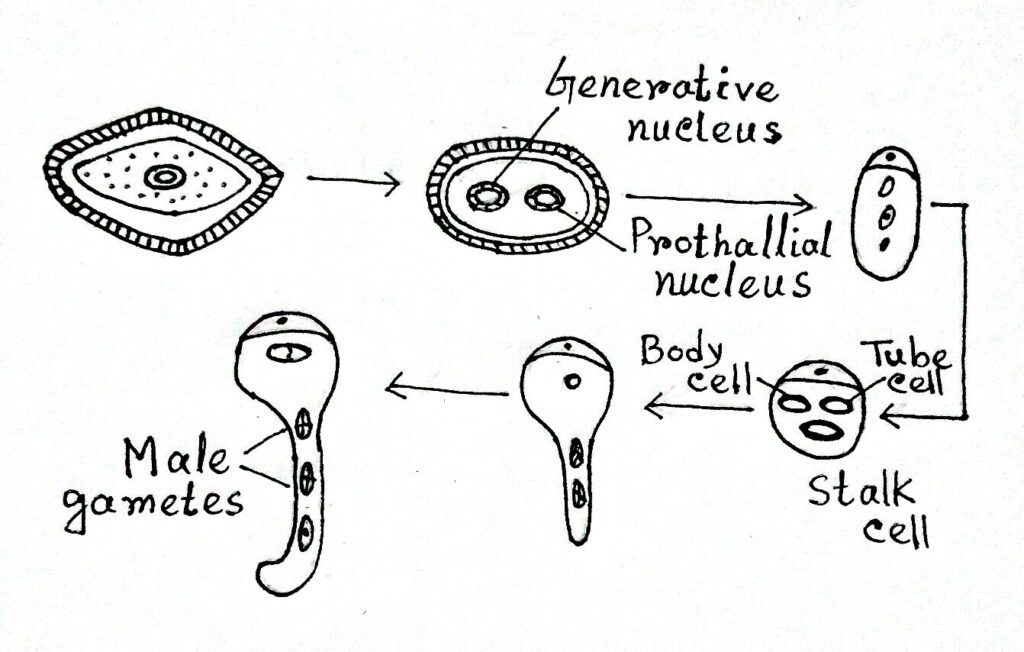
- In Welwitschia, the prothellial cell is not present. So in pollen one generative and one tube cell are present.
- The generative cell divides to form a body cell and a stalk cell. The body cell divides and forms two male gametes.
- In Gnetum, a microspore consists of a prothallial cell, a generative cell, and a tube nucleus.
- The generative cell divides within the pollen tube into two non-motile naked male gametes.
Female Gametophyte
- The functional megaspore is the first cell of the female gametophyte.
- In Ephedra, the megaspore divides by free nuclear division and forms the embryo sac.
- Archegonia are developed from archegonial initial. An archegonium consists of 2 Neck canal cells, one ventral canal cell, and one egg.
- In Gnetum, the megaspore mother cell takes part in the development of the gametophyte. It’s containing four megaspore nuclei and forms a female gametophyte (embryo sac).
- The four megaspores divide continuously and form thousands of female nuclei.
- The embryo sac is tetrasporic, in which 256-1500 nuclei are found.
- At this time cell wall is formed from the chalazal end.
- A few free nuclei in this part become larger and differentiate as eggs.
- Archegonia are not formed in Gnetum female gametophyte.
Fertilization
- In Ephedra, the pollen tube enters the archegonium and releases two gametes.
- The large gamete fuses with the egg and formed a zygote.
- In Welwitschia, the embryo sac moves towards the pollen tube and gets attached to it and the fusion wall disappears.
- Female nuclei enter the pollen tube and fused with male gamete formed zygote.
- In Gnetum, the pollen tube comes in contact with the female gametophyte and the male gamete enters the embryo sac.
- One gamete attaches to the female nucleus and forms a zygote. While the second gamete attaches to the other nucleus and as a result double fertilization occurs.
- At this time, the endosperm is formed due to wall formation at the micropylar end.
Embryo Development
- In Ephedra, the zygote is divided by free nuclear division and forms 8 nuclei.
- Cell wall formation occurs in three to five nuclei and each of the cells formed one embryo. Therefore, polyembryony condition is present in Ephedra but in the end, one embryo gets mature.
- In Welwitschia, the zygote divides and forms a suspensor and an embryonal cell.
- Now the embryonal cell divides periclinally and anticlinally and quadrant and octant are formed. Its marginal cells get enlarged and cover the embryonal cells resulting in a number of embryo formations. But only one embryo matures.
- The embryo consists of two cotyledons, plumules, and hypocotyl.
- In Gnetum, the zygote divides and forms two cells. Both the cells enlarge and form tubes.
- Each tube divides and forms suspensor and embryonal cells.
Seed Germination
- Seeds are endospermic.
- In Ephedra and Welwitschia send germination is epigeal
- But the process of germination of the seed is hypogeal in Gnetum.
Classification of Gnetales
Markgraf (1951) classified order Gnetales into three following families;
- Ephedraceae (one genus- Ephedra)
- Welwitschiaceae (one genus- Welwitschia)
- Gnetaceae (one genus- Gnetum)
