Laminaria is a highly developed parenchymatous marine brown alga of the family Laminariales. It is mostly found in the northern Atlantic and northern Pacific Oceans.
The species of Laminaria are popularly known as “Devil’s aprons” due to their shape. They are also called sea colanders because of the perforations present on the lamina.
| Class: | Phaeophyceae |
| Order: | Laminariales |
| Family: | Laminariaceae |
| Genus: | Laminaria |
Salient Features of Laminaria
The salient features of Laminaria are as follows:
- The plant body is sporophytic and very large in size.
- The sporophyte is externally differentiated into a holdfast, stipe, and blade.
- Internally, the thallus is very complex in structure, and it shows three distinct regions: the meristoderm, cortex, and medulla.
- Growth in Laminaria is intercalary, and the presence of meristematic tissue between the stipe and lamina causes annual replacement of the lamina.
- The sori, which contain sporangia and paraphyses, are developed on both surfaces of the blade.
- Asexual reproduction occurs through the formation of zoospores.
- Zoospores are formed within the sporangium by reduction division (meiosis).
- Gametophytes are heterothallic. On germination, half of the zoospores develop into male gametophytes, and the other half produce female gametophytes.
- Sexual reproduction is of the oogamous type.
- The male sex organ antheridium produces the antherozoid, while the female sex organ oogonium develops into an egg.
- Heteromorphic alternation of generation is present in the life cycle, where the predominant sporophyte alternates with the much reduced gametophyte.

Occurrence of Laminaria
Laminaria is a genus of about 40 species in the order Laminariales (kelp). It occurs exclusively in marine habitats.
The species of Laminaria are found throughout the world, especially along the coasts of the Atlantic and Pacific Oceans. They occur attached to the rocks and stones in the littoral and sublittoral zones of the oceans.
Most of the species are perennial lithophytes, but a few are epiphytes. Some species grow widely in shallow, cold water.
Thallus Structure of Laminaria
The plant body of Laminaria is a diploid sporophyte. It shows a parenchymatous organization (a modification of the filamentous habit).
The plant is very large and may reach up to 2–12 metres or more in height (15 metres in Laminaria farlowii).
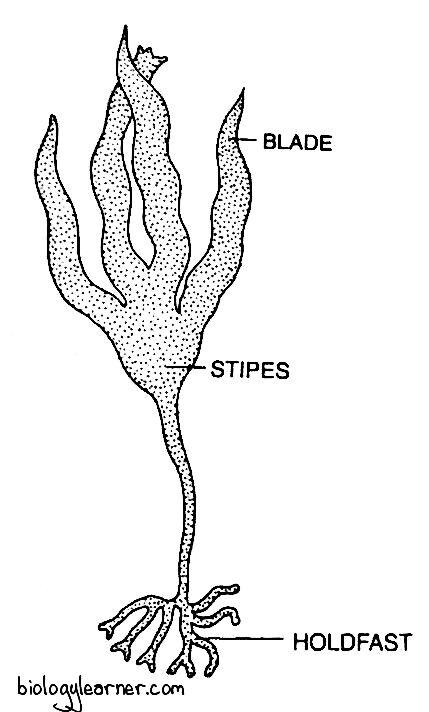
The sporophytic thallus is differentiated externally into three distinct parts: a holdfast, stipe, and blade (lamina).
Holdfast
The holdfast may be a solid disc-like structure. But it usually consists of forked root-like branches called heptera.
Holdfast attaches the plant body to the substratum.
Stipe
The stipe develops on the holdfast. It is always unbranched and is either cylindrical or slightly flattened.
The stipe forms the main axis of the sporophyte and bears terminal blades.
Blade (Lamina)
The blade or lamina may be simple (e.g., L. saccharina) or vertically incised into a number of segments (e.g., L. digitata, L. cloustoni).
The lamina is about 2–4 metres long. It is flat, long, and ribbon-shaped, with a wavy or smooth margin, and has a rough and leathery texture.
Internal Structure of Laminaria
In a transverse section, both the blade and the stipe show three regions: the meristoderm (epidermis), the cortex, and the medulla (central axis). However, the tissues are more clearly marked off in the stipe than in the blade.
Meristoderm (Epidermis)
The meristoderm is the outermost region and is composed of one or two layers of small cubical cells containing chromatophores. It is covered externally by a layer of mucilage.
The meristoderm serves the function of assimilation.
Cortex
The cortex lies between the meristoderm and the central medulla. It is usually differentiated into an outer and an inner cortex.
The outer cortex is much wider and consists of vertically elongated cells with tapering ends, while the inner cortex is composed of longer cells with square ends. The cells formed earlier in the growing seasons are larger. Those developed at the end of the season are smaller. Hence, the cells of the cortex show variations in their sizes.
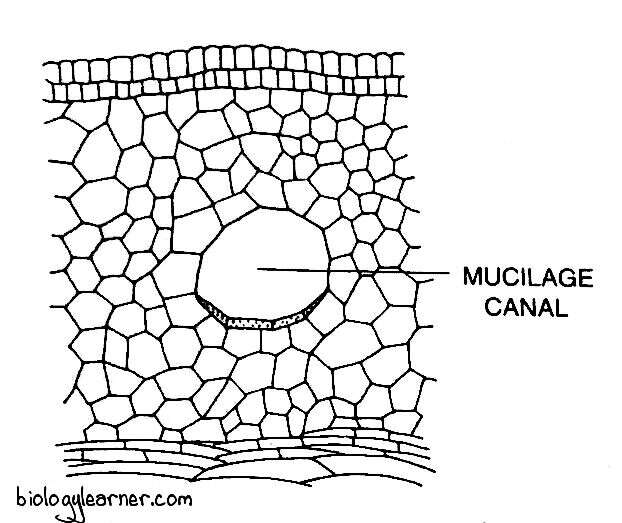
In many species, mucilage ducts can be found in the cortex. The function of cortical cells (cells of the cortex) is to provide storage and mechanical support.
Medulla
The medulla is the innermost layer and is composed of vertical, unbranched filaments (hyphae-like). These hyphae-like filaments lie close to one another in the young plant. But in the mature plant, they lie apart.
The cells are loosely arranged in the filaments. The spaces between these cells are filled with mucilage.

In some filaments of the medulla, the cells elongate vertically but divide infrequently. These vertically elongated cells swell and widen at the transverse walls and form trumpet hyphae. The trumpet hyphae are similar to sieve tubes in vascular plants.
Many pores are present on the transverse walls. These pores have cytoplasmic strands by which the protoplasts of adjoining cells are connected with each other. The wall of the trumpet hypae also possesses some spiral bands of cellulose thickening.
The real function of the trumpet hyphae is still unknown.
Growth of the Thallus
Growth of Laminaria takes place by an intercalary meristem, situated at the junction of the stipe and the blade. The activity of the meristem increases the length of the stipe continuously, but the length of mature blades remains constant.
Reproduction in Laminaria
Laminaria reproduces by all three methods: vegetative, asexual, and sexual.
Vegetative Reproduction
Some species of Laminaria, such as L. longipes and L. sincelarii, reproduce vegetatively by stolons (i.e., special short, slender horizontal branches).
The stolons develop from the holdfast and grow horizontally along the ground surface. Each stolon may give rise to a plantlet.
Asexual Reproduction
Asexual reproduction in Laminaria takes place by the formation of haploid zoospores. Zoospores are developed in the club-shaped unilocular (consisting of a single cavity) sporangia.
During the summer or early autumn, the sporangia are borne in widely extensive sori on both surfaces of the blade and nearly cover the entire area. Many hair-like cells called paraphyses are present intermingled with the sporangia.
Development of Sporangium
In Laminaria, the sporangia develop from the superficial cells (outer epidermal cells) of the blade. Initially, the superficial cell enlarges and divides transversely into an upper cell and a lower basal cell.
The upper cell functions as the paraphysis initial. It elongates and forms the paraphysis (an elongated thread with a club-shaped upper region). The upper region of the paraphysis contains many chromatophores and fucosan vesicles.

Each paraphysis is covered by a prominent mucilage cap at the top. The mucilage caps of all the paraphyses in a sorus remain attached firmly together, keeping the sorus intact.
Simultaneously, by the side of the paraphyses, the basal cells enlarge and develop into sporangia. The sporangia remain between the paraphyses.
The nucleus of the sporangium first divides meiotically, followed by several successive mitotic divisions, resulting in the formation of 32–64 haploid daughter nuclei. Each nucleus, along with some cytoplasm, is ultimately metamorphosed into a single biflagellate zoospore.
Zoospores
The zoospore is a biflagellate, pear-shaped structure without an eye spot (Henry and Cole, 1982). The two flagella are inserted laterally and are of unequal length. The longer flagellum is of the tinsel type, while the shorter one is of the whiplash type.
The mature sporangium dehisces by the lateral pressure of paraphyses on the sporangium. The apex of the sporangium degenerates, and mature zoospores are liberated from the sporangium. The liberated zoospores remain enclosed in a watery gelatinous sheath, which persists for a minute or more and finally dissolves, leaving the zoospores swimming freely in the water.
Germination of Zoospores
About half of the zoospores germinate into male gametophytic plants and the other half into female gametophytic plants.
After swimming for some time, the zoospores settle down to germinate. Each zoospore withdraws its flagella, becomes rounded, and secretes a wall. It develops into the germ tube (a small protuberance) on germination.
The germ tube increases in length, and the nucleus and chromatophores move into its apex. The apex becomes bulbous and appears almost like a dumbbell.
The bulbous region of the germ tube develops into a small filamentous gametophyte.
Sexual Reproduction
Sexual reproduction in Laminaria is of the oogamous type. The male gametes, called antherozoides, are produced in antheridium, and the female gamete, known as an egg, is produced in oogonium.
The species of Laminaria are heterothallic, i.e., dioecious. Antheridia and oogonia are borne on different gametophytic plants.
Antheridium
The male gametophyte is a many-celled, simple, filamentous, and branched structure. It produces antheridia.
The antheridia develop singly or in groups at the tips of the lateral branches of many-celled male gametophytes. Each antheridium is small, unicellular, and spherical in shape.
The protoplast of each antheridium metamorphoses into a single pyriform or ellipsoidal antherozoid with two unequal, laterally inserted flagella.
Antherozoids are liberated through the apical portion of the antheridia. After the liberation of the antherozoid, the male plant degenerates.
Oogonium
The female gametophyte is sparingly branched. It develops oogonia at the 2-3 celled stage. The oogonia may be terminal or intercalary, and they are longer and thicker than the other cells of the female gametophyte.
The protoplast of the oogonium develops into an egg. At maturity, the egg is extruded through the apical pore of the oogonium but remains attached to the apex of the oogonial wall.
Fertilization
The liberated antherozoid swims in water. It comes to the oogonium and fuses with the egg to form the diploid zygote.
Germination of Zygote
After fertilization, the zygote secretes a thin membrane around itself to form an oospore. The oospore, without passing through any resting period, develops into a new sporophytic plant.
The diploid zygote, i.e., the oospore, undergoes several successive transverse divisions and forms a vertical row of 4–10 cells. Except for the lowermost cell, all other cells further divide to form an expanded, thin, and flat blade.
The lowermost cell elongates and forms the first rhizoid. Later, many branched rhizoids are developed.
The lower part of the blade divides further, resulting in the formation of the meristematic region. With further development, the lower portion of the meristem gets differentiated into the stipe.
