Ectocarpus is a filamentous heterotrichous marine brown alga of the family Ectocarpaceae. It is the most primitive genus of brown algae.
Due to the small mature thallus and relatively quick completion of the life cycle, Ectocarpus serves as a model organism for the study of the genomics of multicellularity (multicellular organisms).
| Class: | Phaeophyceae |
| Sub-Class: | Phaeophycidae |
| Order: | Ectocarpales |
| Family: | Ectocarpaceae |
| Genus: | Ectocarpus |
Salient Features of Ectocarpus
The salient features of Ectocarpus are as follows:
- The plant body is brown, much branched, and filamentous.
- The filaments are heterotrichous in nature and are differentiated into a prostrate system and an erect system.
- Both gametophytic (haploid) and sporophytic (diploid) plants are isomorphic, i.e., morphologically alike.
- The sporophytic plant bears both unilocular and plurilocular sporangia, while the gametophytic plant consists only of plurilocular gametangia (similar to plurilocular sporangia).
- Asexual reproduction occurs through the formation of biflagellate zoospores within the unilocular and plurilocular sporangia.
- The unilocular sporangium produces haploid zoospores (n), which, on germination, develop into new gametophytic plants.
- In plurilocular sporangium, diploid zoospores (2n) are formed. Diploid zoospores, on germination, give rise to sporophytic plants.
- Sexual reproduction is of the isogamous, anisogamous (clump-formation), or oogamous type.
- Gametes are produced in plurilocular gametangia.
- After fertilization, the diploid zygote germinates directly into a new diploid sporophytic plant.
- Presence of a distinct isomorphic alternation of generation in the life cycle.

Occurrence of Ectocarpus
Ectocarpus is a genus of about 410 species. It is found all over the world, mainly in the colder seas of temperate and tropical regions.
All the species of Ectocarpus occur in marine habitats either as free-floating, epiphytes or lithopytes. Most of the species are lithophytes and grow on rocks in the littoral and sublittoral zones of the oceans. E. coniferus and E. breviarticulatus occur as epiphytes on larger algae like Laminaria and Fucus. E. arabicus is found epiphytically on the leaves of Sargassum tenerrimum.
Ectocarpus fasciculatus is an epizoic species, grows on the fins of certain fish, while E. dermonematis is endophytic. Species like E. carver and E. spongiosus are free-floating.
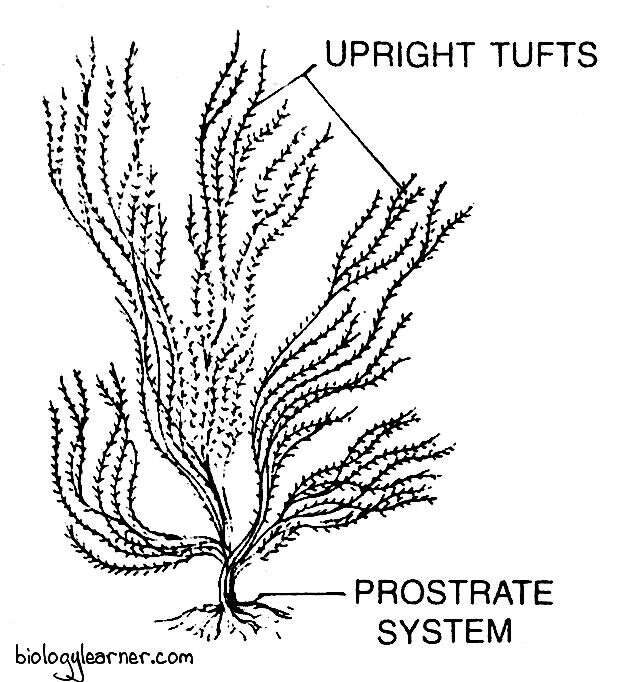
Thallus Structure of Ectocarpus
The plant body of Ectocarpus is multicellular, uniseriate, branched, filamentous and brown or yellow in color. It shows a heterotrichous habit, which is differentiated into a prostrate system and an erect system.
Prostrate or Creeping System
The prostrate system is profusely branched and creeps over the substratum. It is firmly attached to the substratum with the help of rhizoids.
In free-floating species, the prostrate system is poorly developed.
Erect or Projecting System
The erect system arises from the prostrate system. It consists of well-developed, branched filaments, which remain free-floating in the water.
Each filament is several inches long and monosiphonous, i.e., uniseriate. But the lower part becomes polysiphonous (multiseriate) due to longitudinal division (e.g., E. geminifructus).
The main axis of the thallus bears lateral branches. The branches arise very close to the septa (e.g., E. arabicus, E. enhali). The apical portion of each branch terminates into an acute point to form an elongated, hyaline, hair-like trichome.
In some species, the older portions of the main axis may be surrounded by a layer of descending rhizoid-like branches and become corticated.
Cell Structure of Ectocarpus
The cells of the filament are rectangular or cylindrical, placed one above the other in a single series.
Each cell is surrounded by a thick,bi-layered cell wall. The cell wall is differentiated into an inner cellulosic layer and an outer pectic layer. The characteristic gelatinous substance composed of algin and fucoidan is also present in the cell wall.
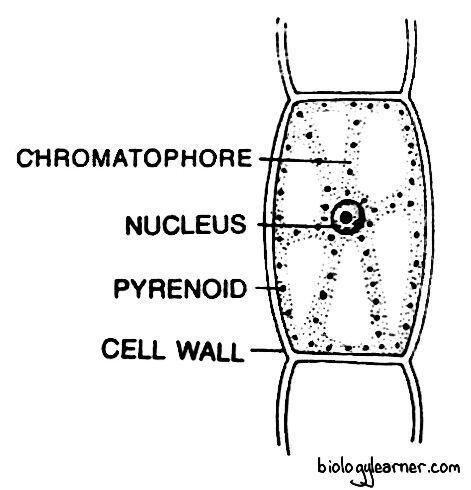
A thin cell membrane encloses the proplast. The protoplast contains a single nucleus and many chromatophores with pyrenoid-like bodies. The chromatophores are variable in number and shape in different species. They may be discoid or ribbon-shaped.
Many pigments, such as chlorophyll-a, chlorophyll-c, β-carotene, and fucoxanthin, are present in the chromatophores. The pigment fucoxanthin gives Ectocarpus its characteristic brown colour.

Each cell possesses many large vacuoles in the cytoplasm called physodes. The physodes contain polyphenols, which are thought to be lysosomal in function.
The reserve food material is in the form of oil, such as mannitol and laminarin, found in the cytoplasm.
Growth of the Thallus
Growth of Ectocarpus is apical in the prostrate filaments. But the erect filaments show intercalary, diffuse, or trichothallic growth due to the intercalary meristem.
An intercalary meristem (short meristematic cells) called the trichothallic meristem is present at the base of each erect branch. The cells of the meristem cut off segments at both ends.
The cells developed above increase the length of the terminal hair, while the lower cells increase the length of the vegetative cells of the branch.
Reproduction in Ectocarpus
In the life cycle of Ectocarpus, two types of thalli are found: the diploid sporophyte and the haploid gametophyte. Asexual reproduction takes place in the sporophytic plants, while sexual reproduction occurs in the gametophytic plants.
Thus, Ectocarpus reproduces both asexually and sexually.
Asexual Reproduction
Asexual reproduction in Ectocarpus occurs in the sporophytic plants through the formation of biflagellate zoospores. Zoospores are haploid as well as diploid and are developed in the sporangia.
Sporangia are of two types: unilocular sporangia (one-celled) and plurilocular sporangia (many-celled). The unilocular sporangia form haploid zoospores, while the plurilocular sporangia produce diploid zoospores. Both the unilocular and plurilocular sporangia are present on the same sporophytic plant.
Unilocular Sporangia
The unilocular sporangium is a stalked, globular, or pear-shaped structure. It is one-celled, consisting of dense cytoplasm with many chromatophores.
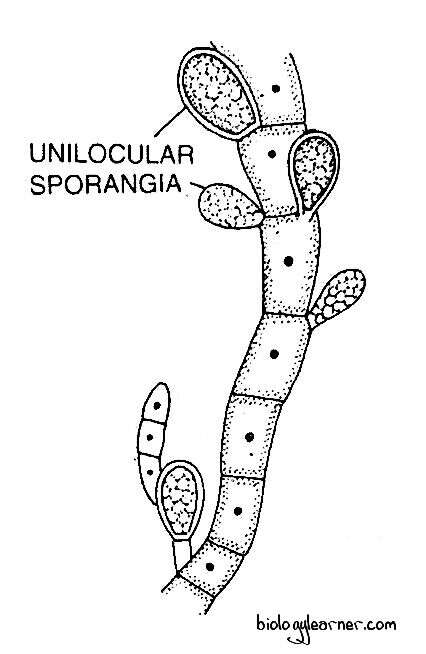
The unilocular sporangia develop from the terminal cells of the short lateral branches. The terminal cell enlarges in size and functions as the sporangial initial.
The cytoplasm and the number of chromatophores also increase, and the sporangial initial becomes globose or ellipsoidal.
The diploid nucleus of the initial cell first undergoes meiosis, followed by several successive mitotic divisions, resulting in the formation of 32–64 haploid daughter nuclei.
Each nucleus accumulates some cytoplasm and ultimately metamorphoses into a single biflagellate zoospore, called the meiozoospore or gonozoospore.
The zoospore or meiozoospore is a haploid, biflagellate, pear-shaped structure. It contains a single nucleus, an eye spot, and a chromatophore. The two flagella are unequal and inserted laterally. The anterior flagellum is longer and of the tinsel type (i.e., pantonematic), directed forward, while the posterior one is short and of the whiplash type (i.e., acronematic), directed backward.
During the liberation of zoospores, the apex of the sporangial wall degenerates. The mature zoospores are liberated from the sporangium in a gelatinous mass. After some time (i.e., 30–60 seconds), they get free from the gelatinous mass and swim freely in the water.
Each zoospore swims for about 30 minutes or more. Then it settles down on the substratum by its anterior end. The zoospore withdraws its flagella, becomes rounded, and forms a wall. It develops into a short germ tube, which becomes separated by a septum.
The germ tube divides many times and forms a prostrate filament. A few cells of the prostrate filament develop erect filaments.
In this way, the meiozoospores germinate into the haploid gametophytic plants.
Plurilocular Sporangium
The plurilocular sporangium is an elongated, multicellular, cone-like structure developed on the diploid sporophytic thallus. It may be sessile or stalked.
Similar to the unilocular sporangia, the plurilocular sporangia also develop initially at the terminal cells of the short lateral branches. The terminal cell functions as the sporangial initial. It enlarges, accumulates many choramtophores, and becomes spherical.
The sporangial initial cell now divides by repeated transverse mitotic divisions and forms a vertical row of 6–12 cells. The cells then undergo several vertical divisions. As a result, an elongated, multi-chambered, cone-like structure is formed, called the plurilocular sporangium.
The plurilocular sporangium contains many small cubical cells arranged in 20–40 transverse tiers. The protoplast of each cell directly metamorphoses into a single uninucleate, biflagellate zoospore. Since there is no reduction division during the formation of zoospores in the plurilocular sporangium, the zoospores are diploid.
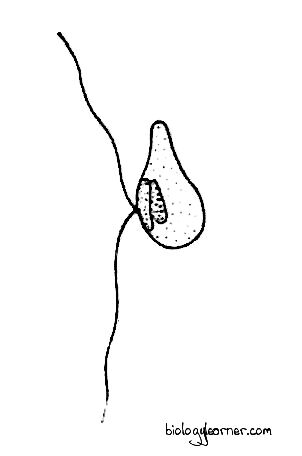
The zoospores developed from the plurilocular sporangia are structurally identical to the meiozoospores (haploid zoospores) produced in the unilocular sporangia.

The zoospores are liberated gradually through a small apical or lateral aperture developed on the wall of the sporangium. Later, each of the zoospores gives rise to a new diploid sporophytic plant.
The plurilocular sporangia play no role in the alternation of generation, as the diploid zoospores are produced for the reduplication of sporophytic generation. Thus, these diploid zoospores are also called neutral spores, and the plurilocular sporangia are also known as neutral sporangia.
Sexual Reproduction
Sexual reproduction in Ectocarpus is of both the isogamous and anisogamous types. Anisogamy is of two types: morphological anisogamy and physiological anisogamy.
Most of the species are isogamous and monoecious (homothallic), i.e., both types of gametes (male and female) are produced in the same plant (e.g., E. pusillus, E. globifer). Some species are anisogamous and dioecious (heterothallic), i.e., male and female gametes come from different plants (e.g., E. secundus, E. siliculosus).
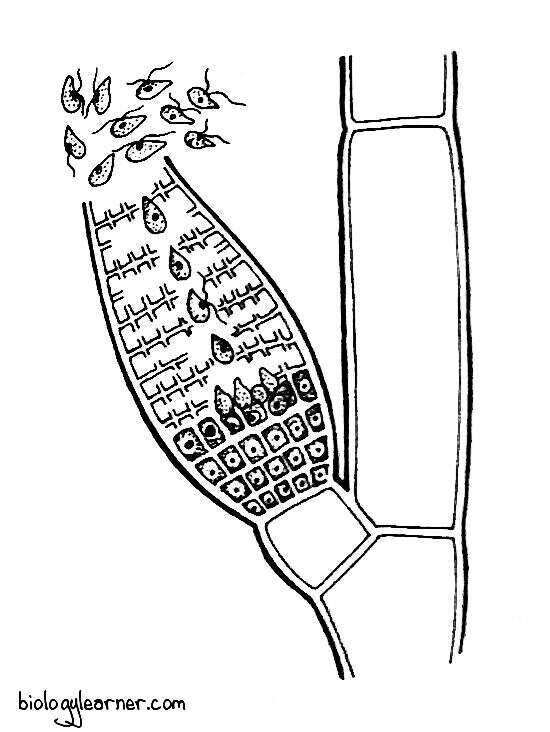
The gametes are produced in the large, elongated sex organs known as the plurilocular gametangia.

Plurilocular Gametangium
The plurilocular gametangium is a large, elongated, multi-chambered, sessile or short-stalked conical structure. They morphologically resemble the plurilocular sporangia but produce haploid gametes.
The gametangium also develops in a similar way to the plurilocular sporangium, at the tip of a short lateral branch. They consists of numerous small cubical cells.
Later on, the protoplast of each cell metamorphoses into a single haploid gamete. The gametes are structurally identical to zoospores produced in asexual reproduction, but they are relatively smaller in size.
Liberation of gametes occurs through a terminal or lateral pore. After the liberation of gametes, gametic fusion takes place.
Isogamy
Isogamy can be seen in species like E. pusillus and E. globifer (Goebel, 1878). In this type, the fusion takes place between the morphologically identical gametes.

The gametes are similar in size, shape, and structure. They come from the same plant, or even the same gametangium.
After fusion between two gametes, a diploid zygote is formed.
Morphological Anisogamy
Ectocarpus secundus shows morphological anisogamy (Bornet, 1891; Sauvageau, 1896). Here, two fusing gametes are dissimilar in size and are formed in different gametangia.
The smaller male gametes, or microgametes, are developed in the microgametangium (i.e., male gametangium), while the larger female gametes, or macrogametes, are produced in the megagametangium (i.e., female gametangium).
After liberation, the microgamete and the macrogamete come close to each other. Now both gametes fuse together and form a diploid zygote.
Physiological Anisogamy (Clump Formation)
Physiological anisogamy is observed in E. siliculosus (Berthold, 1881).
The fusing gametes are morphologically alike but dissimilar in sexual behaviour. Some gametes are less active and are called female gametes, while other gametes are more active and are known as male gametes.
The female gamete becomes passive and soon comes to rest on a substratum. Many active male gametes cluster around the surface of the female gamete by their forwardly directed long flagella. This phenomenon is known as clump formation.

Finally, one of the male gametes fuses with the female gamete. The remaining active gametes swim away, and they gradually get destroyed. After fusion (plasmogamy and karyogamy), a diploid zygote is formed.

Germination of Zygote
The diploid zygote, without passing through any resting stage, develops into a new diploid sporophytic plant. During the germination of the zygote, no reduction division (zygotic meiosis) occurs.
